Значение скелетной мускулатуры. Режимы мышечных сокращений. Классификация упрощает изучение
Скелетная мышечная ткань
Схема скелетной мышцы в разрезе.

Строение скелетной мышцы
Скелетная (поперечно-полосатая) мышечная ткань - упругая, эластичная ткань , способная сокращаться под влиянием нервных импульсов : один из типов мышечной ткани . Образует скелетную мускулатуру человека и животных, предназначенную для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания. Мышцы состоят на 70-75 % из воды.
Гистогенез
Источником развития скелетной мускулатуры являются клетки миотомов - миобласты. Часть из них дифференцируется в местах образования так называемых аутохтонных мышц. Прочие же мигрируют из миотомов в мезенхиму ; при этом они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела. В ходе дифференцировки возникает 2 клеточные линии. Клетки первой сливаются, образуя симпласты - мышечные трубки (миотубы). Клетки второй группы остаются самостоятельными и дифференцируются в миосателлиты (миосателлитоциты).
В первой группе происходит дифференцировка специфических органелл миофибрилл , постепенно они занимают большую часть просвета миотубы, оттесняя ядра клеток к периферии.
Клетки второй группы остаются самостоятельными и располагаются на поверхности мышечных трубок.
Строение
Структурной единицей мышечной ткани является мышечное волокно. Оно состоит из миосимпласта и миосателлитоцитов (клеток-спутниц), покрытых общей базальной мембраной .
Длина мышечного волокна может достигать нескольких сантиметров при толщине в 50-100 микрометров.
Строение миосимпласта
Строение миосателлитов
Миосателлиты - одноядерные клетки, прилежащие к поверхности миосимпласта. Эти клетки отличаются низкой дифференцировкой и служат взрослыми стволовыми клетками мышечной ткани. В случае повреждения волокна или длительном увеличении нагрузки клетки начинают делиться, обеспечивая рост миосимпласта.
Механизм действия
Функциональной единицей скелетной мышцы является моторная единица (МЕ). МЕ включает в себя группу мышечных волокон и иннервирующий их мотонейрон . Число мышечных волокон, входящих в состав одной МЕ, варьирует в разных мышцах. Например, там, где требуется тонкий контроль движений (в пальцах или в мышцах глаза), Моторные единицы небольшие, они содержат не более 30 волокон. А в икроножной мышце, где тонкий контроль не нужен, в МЕ насчитывается более 1000 мышечных волокон.
Моторные единицы одной мышцы могут быть разными. В зависимости от скорости сокращения моторные единицы разделяют на медленные (slow (S-МЕ)) и быстрые (fast (F-МЕ)). А F-МЕ в свою очередь делят по устойчивости к утомлению на устойчивые к утомлению (fast-fatigue-resistant (FR-МЕ)) и быстроутомляемые (fast-fatigable (FF-МЕ)).
Соответствующим образом подразделяют иннервирующие данные МЕ мотонейроны. Существуют S-мотонейроны (S-МН), FF-мотонейроны (F-МН) и FR -мотонейроны (FR-МН) S-МЕ характеризуются высоким содержанием белка миоглобина, который способен связывать кислород (О2). Мышцы, преимущественно состоящие из МЕ этого типа, за их темно-красный цвет называются красными. Красные мышцы выполняют функцию поддержания позы человека. Предельное утомление таких мышц наступает очень медленно, а восстановление функций происходит наоборот, очень быстро.
Такая способность обуславливается наличием миоглобина и большого числа митохондрий . МЕ красных мышц, как правило, содержат большое количество мышечных волокон. FR-МЕ составляют мышцы, способные выполнять быстрые сокращения без заметного утомления. Волокна FR-ME содержат большое количество митохондрий и способны образовывать АТФ путем окислительного фосфорилирования.
Как правило, число волокон в FR-ME меньше, чем в S-ME. Волокна FF-ME характеризуются меньшим содержанием митохондрий, чем в FR-ME, а также тем, что АТФ в них образуется за счет гликолиза . В них отсутствует миоглобин , поэтому мышцы, состоящие из МЕ этого типа, называют белыми. Белые мышцы развивают сильное и быстрое сокращение, но довольно быстро утомляются.
Функция
Данный вид мышечной ткани обеспечивает возможность выполнения произвольных движений. Сокращающаяся мышца воздействует на кости или кожу, к которым она прикрепляется. При этом один из пунктов прикрепления остаётся неподвижным - так называемая точка фиксации (лат. púnctum fíxsum ), которая в большинстве случаев рассматривается в качестве начального участка мышцы. Перемещающийся фрагмент мышцы называют подвижной точкой , (лат. púnctum móbile ), которая является местом её прикрепления. Тем не менее, в зависимости от выполняемой функции, punctum fixum может выступать в качестве punctum mobile , и наоборот.
Примечания
См. также
Литература
- Ю.И. Афанасьев, Н.А. Юрина, Е.Ф. Котовский Гистология. - 5-е изд., перераб. и доп.. - Москва: Медицина, 2002. - 744 с. - ISBN 5-225-04523-5
Ссылки
- - Механизмы развития мышечной ткани (англ.)
| Мышечная система | |
|---|---|
Wikimedia Foundation . 2010 .
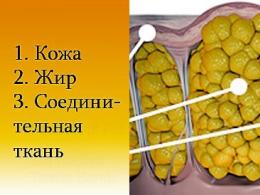
Скелетная (соматическая) мускулатура представлена большим количеством (более 200) мышц. Каждая мышца имеет опорную часть - соединительнотканную строму и рабочую часть - мышечную паренхиму. Чем большую статическую нагрузку выполняет мышца, тем больше развита в ней строма.
Снаружи мускул одет соединительнотканной оболочкой, которая называется наружным перимизием - perimysium. На различных мышцах он разной толщины. От наружного перимизия внутрь отходят соединительнотканные перегородки - внутренний перимизий, окружающий мышечные пучки различной величины. Чем большую статическую функцию несет мышца, тем более мощные соединительнотканные перегородки в ней расположены, тем их больше. На внутренних перегородках в мышцах могут закрепляться мышечные волокна, проходят сосуды и нервы. Между мышечными волокнами проходят очень нежные и тонкие соединительнотканные прослойки, называемые эндомизием - endomysium.

В этой строме мышцы, представленной наружным и внутренним перимизием и эндомизием, закономерно упакована мышечная ткань (мышечные волокна, образующие мышечные пучки), формирующая различной формы и величины мышечное брюшко. Строма мышцы по концам мышечного брюшка образует сплошные сухожилия, форма которых зависит от формы мышц. Если сухожилие шнурообразно, оно называется просто сухожилием - tendo. Если сухожилие плоское, идет от плоского мускульного брюшка, то оно называется апоневрозом.
В сухожилии также различают наружные и внутренние оболочки (мезотендиний - mesotendineum). Сухожилия очень плотны, компактны, образуют прочные шнуры, обладающие большой сопротивляемостью на разрыв. Коллагеновые волокна и пучки в них расположены строго продольно, благодаря чему сухожилия становятся менее утомляемой частью мышцы. Закрепляются сухожилия на костях, проникая в толщу костной ткани в виде шарпеевских волокон (связь с костью настолько крепка, что скорее разорвется сухожилие, чем оно оторвется от кости). Сухожилия могут переходить на поверхность мышцы и покрывать их на большем или меньшем расстоянии, образуя блестящую оболочку, которая называется сухожильным зеркалом.

В определенных участках в мышцу входят сосуды, ее кровоснабжающие, и нервы, ее иннервирующие. Место вступления их называется воротами органа. Внутри мышцы сосуды и нервы разветвляются по внутреннему перимизию и доходят до его рабочих единиц - мышечных волокон, на которые сосуды образуют сети капилляров, а нервы разветвляются на:
1) чувствительные волокна - идут от чувствительных нервных окончаний проприорецепторов, расположенных" во всех участках мышц и сухожилий, и выносят импульс, направляющийся через клетку спинального ганглия в мозг;
2) двигательные нервные волокна, проводящие импульс от мозга: а) к мышечным волокнам, заканчиваются на каждом мышечном волокне особой моторной бляшкой, б) к сосудам мышц - симпатические волокна, несущие импульс от мозга через клетку симпатического ганглия к гладким мышцам сосудов, в) трофические волокна, заканчивающиеся на соединительнотканной основе мышцы.
Поскольку рабочей единицей мышц является мышечное волокно, то именно их количество определяет силу мышцы; не от длины мышечных волокон, а от количества их в мышце зависит сила мышцы. Чем больше мышечных волокон в мышце, тем она сильнее. Длина мышечных волокон обычно не превышает 12-15 см, подъемная сила мышцы в среднем равна 8-10 кг на 1 см 2 физиологического поперечника. При сокращении мышца укорачивается на половину своей длины. Чтобы подсчитать количество мышечных волокон, делают разрез перпендикулярно их продольной, оси; полученная площадь поперечно перерезанных волокон - это физиологическими поперечник. Площадь разреза всей мышцы перпендикулярная ее продольной оси называется анатомическим поперечником. В одной и той же мышце может быть один анатомический и несколько физиологических поперечников, образовавшихся в том случае, если в мышце мышечные волокна короткие и имеют различное направление. Так как сила мышцы зависит от количества мышечных волокон в них, то она выражается отношением анатомического поперечника к физиологическому. В мышечном брюшке имеется всего один анатомический поперечник, а физиологических может быть различное количество (1:2, 1:3,..., 1:10 и т.д.). Большое количество физиологических поперечников свидетельствует о силе мышцы.
Мышцы бывают светлые и темные. Цвет их зависит от функции, строения и кровенаполнения. Темные мышцы богаты миоглобином (миогематином) и саркоплазмой, они более выносливые. Светлые мышцы беднее этими элементами, они более сильные, но менее выносливые. У разных животных, в различном возрасте и даже в разных участках тела цвет мышц бывает различен: самые темные они у лошади, гораздо светлее у свиней; у молодняка светлее, чему взрослых; на конечностях темнее, чем на теле; у диких животных темнее, чем у домашних; у кур грудные мышцы белые, у диких птиц темные.
Структурно-функциональной единицей скелетной мышцы является симпласт или мышечное волокно - огромная клетка, имеющая форму протяженного цилиндра с заостренными краями (под наименованием симпласт, мышечное волокно, мышечная клетка следует понимать один и тот же объект).
Длина мышечной клетки чаще всего соответствует длине целой мышцы и достигает 14 см, а диаметр равен нескольким сотым долям миллиметра. скелетный мышца строение развитие
Мышечное волокно, как и любая клетка, окружено оболочкой - сарколемой. Снаружи отдельные мышечные волокна окружены рыхлой соединительной тканью, которая содержит кровеносные и лимфатические сосуды, а так же нервные волокна.
Группы мышечных волокон, образуют пучки, которые, в свою очередь, объединяются в целую мышцу, помещенную в плотный чехол соединительной ткани переходящей на концах мышцы в сухожилия, крепящиеся к кости (рис.1).
Рис. 1.
Усилие, вызываемое сокращением длины мышечного волокна, передается через сухожилия костям скелета и приводит их в движение.
Управление сократительной активностью мышцы осуществляется с помощью большого числа мотонейронов (рис. 2) - нервных клеток, тела которых лежат в спинном мозге, а длинные ответвления - аксоны в составе двигательного нерва подходят к мышце. Войдя в мышцу, аксон разветвляется на множество веточек, каждая из которых подведена к отдельному волокну.

Рис. 2.
Таким образом, один мотонейрон иннервирует целую группу волокон (так называемая нейромоторная единица), которая работает как единое целое.
Мышца состоит из множества нервно моторных единиц и способна работать не всей своей массой, а частями, что позволяет регулировать силу и скорость сокращения.
Для понимания механизма сокращения мышцы необходимо рассмотреть внутреннее строение мышечного волокна, которое, как вы уже поняли, сильно отличается от обычной клетки. Начнем с того, что мышечное волокно многоядерно. Связано это с особенностями формирования волокна при развитии плода. Симпласты (мышечные волокна) образуются на этапе эмбрионального развития организма из клеток предшественников - миобластов.
Миобласты (неоформленные мышечные клетки) интенсивно делятся, сливаются и образуют мышечные трубочки с центральным расположением ядер. Затем в мышечных трубочках начинается синтез миофибрилл (сократительных структур клетки см. ниже), и завершается формирование волокна миграцией ядер на периферию. Ядра мышечного волокна к этому времени уже теряют способность к делению, и за ними остается только функция генерации информации для синтеза белка.
Но не все миобласты идут по пути слияния, часть из них обособляется в виде клеток-сателлитов, располагающихся на поверхности мышечного волокна, а именно в сарколеме, между плазмолемой и базальной мембраной - составными частями сарколемы. Клетки-сателлиты, в отличие от мышечных волокон, не утрачивают способность к делению на протяжении всей жизни, что обеспечивает увеличение мышечной массы волокон и их обновление. Восстановление мышечных волокон при повреждении мышцы возможно благодаря клеткам-сателлитам. При гибели волокна, скрывающиеся в его оболочке, клетки-сателиты активизируются, делятся и преобразуются в миобласты.
Миобласты сливаются друг с другом и образуют новые мышечные волокна, в которых затем начинается сборка миофибрилл. То есть при регенерации полностью повторяются события эмбрионального (внутриутробного) развития мышцы.
Помимо многоядерности отличительной чертой мышечного волокна является наличие в цитоплазме (в мышечном волокне ее принято называть саркоплазмой) тонких волоконец - миофибрилл (рис.1), расположенных вдоль клетки и уложенных параллельно друг другу. Число миофибрилл в волокне достигает двух тысяч.
Миофибриллы являются сократительными элементами клетки и обладают способностью уменьшать свою длину при поступлении нервного импульса, стягивая тем самым мышечное волокно. Под микроскопом видно, что миофибрилла имеет поперечную исчерченность - чередующиеся темные и светлые полосы.
При сокращении миофибриллы светлые участки уменьшают свою длину и при полном сокращении исчезают вовсе. Для объяснения механизма сокращения миофибриллы около пятидесяти лет назад Хью Хаксли была разработана модель скользящих нитей, затем она нашла подтверждение в экспериментах и сейчас является общепринятой.
К основным функциональным свойствам мышечной ткани относятся возбудимость, сократимость, растяжимость, эластичность и пластичность.
Возбудимость - способность мышечной ткани приходить в состояние возбуждения при действии тех или иных раздражителей. В обычных условиях происходит электрическое возбуждение мышцы, вызываемое разрядом мотонейронов в области концевых пластинок. Возникающий под влиянием медиатора потенциал концевой пластинки (ПКП), достигнув порогового уровня (около 30 мВ), вызывает генерацию потенциала действия, распространяющегося в обе стороны мышечного волокна.
Возбудимость мышечных волокон ниже возбудимости нервных волокон, иннервирующих мышцы, хотя критический уровень деполяризации мембран в обоих случаях одинаков. Это объясняется тем, что потенциал покоя мышечных волокон выше (около 90 мВ) потенциала покоя нервных волокон (70 мВ). Следовательно, для возникновения потенциала действия в мышечном волокне необходимо деполяризовать мембрану на большую величину, чем в нервном волокне.
Способность мышцы реагировать на раздражение ее двигательного мотонейрона, т.е. на импульсы, приходящие к ней по нерву, обозначается как непрямая возбудимость мышцы. Однако возбудимостью обладает и само мышечное волокно. Это доказывается раздражением участков мышцы, где отсутствуют окончания двигательного нерва.
Можно исключить влияние нервных элементов на мышцу, подвергнув ее отравлению некоторыми ядами (например, кураре). В этом случае возбуждение с нерва на мышцу не передается, но нерв и мышца сами по себе продолжают функционировать, т.е. мышца продолжает реагировать на непосредственно наносимое на нее раздражение. Таким образом, опыты подобного рода с несомненностью устанавливают наличие в мышечном волокне так называемой прямой возбудимости, т.е. способности мышечных волокон реагировать и на раздражение, действующее непосредственно и на них, а не через нервные волокна.
И прямая и непрямая возбудимость мышцы обусловлена функцией мембраны мышечного волокна. Возбуждение в мышцах проводится изолированно, т.е. не переходит с одного мышечного волокна на другое. Скорость распространения возбуждения в белых и красных волокнах скелетных мышц различна: в белых волокнах она равна 12 - 15, в красных - 3 - 4 м/с.
В мышцах имеется пассивный упругий компонент, который включает сухожилия, соединительную ткань, покрывающую мышечные волокна, их пучки и мышцу в целом, а также упругие образования боковых поперечных мостиков миозиновой нити. Поэтому скелетная мышца - упругое образование. Упругостью обладают активные сократительные и пассивные компоненты мышцы, которые и обеспечивают растяжимость, эластичность и пластичность мышц.
Растяжимость - свойство мышцы удлиняться под влиянием силы тяжести (нагрузки). Чем больше нагрузка, тем больше растяжимость мышцы. Растяжимость зависит и от вида мышечных волокон. Красные волокна растягиваются больше, чем белые, мышцы с параллельными волокнами удлиняются больше, чем перистые. Даже в условиях покоя мышцы всегда несколько растянуты, поэтому они упруго напряжены (находятся в состоянии мышечного тонуса).
Эластичность - свойство деформированного тела возвращаться к первоначальному своему состоянию после удаления силы, вызвавшей деформацию. Это свойство изучается при растяжении мышцы грузом. После удаления груза, мышца не всегда достигает первоначальной длины, особенно при длительном растяжении или под действием большого груза. Это связано с тем, что мышца теряет свойство совершенной упругости.
Пластичность - (греч. Plastikos - годный для лепки, податливый) свойство тела деформироваться под действием механических нагрузок, сохранять приданную или длину или вообще форму после прекращения действия внешней деформирующей силы. Чем длительнее действует большая внешняя сила, тем сильнее пластические изменения.
Пластичность мышц связана и с остаточным укорочением мышц после длительного тетанического сокращения, или контрактуры. Красные волокна, которые удерживают тело в определенном положении, обладают большей пластичностью, чем белые.
При прямом или непрямом раздражении мышца укорачивается или же развивает напряжение в продольном направлении. Это изменение формы или напряжения мышцы носит название мышечного сокращения, следовательно, сократимость - это специфическая деятельность мышечной ткани при ее возбуждении.
Для изучения свойств мышц в учебных целях и в эксперименте в качестве объекта обычно используют нервно-мышечный препарат лягушки, а в качестве раздражителя - электрический ток. Запись сокращений мышцы на приборе миографе при прямом или непрямом раздражении называется миографией. Скорость и сила ответной реакции скелетной мышцы на раздражение зависит не только от параметров раздражителя, но и от типа мышечных волокон. Сократимость и возбудимость мышц разного вида различна.
По скорости сокращения различают быстрые и медленные мышечные волокна. В быстрых волокнах обычно лучше развит саркоплазматический ретикулум, они слабее снабжены кровеносными сосудами, имеют более крупные и длинные волокна, их расслабление после сокращения происходит в 50 - 100 раз быстрее, чем медленных волокон. Организм для выполнения статической работы (например, поддержание позы) использует главным образом медленные, тонические красные мышцы, а для скоростных движений - быстрые белые мышцы.
Различают различные режимы сокращения мышц, которые определяются частотой и силой поступающих импульсов возбуждения.
На прямые и непрямые раздражения частотой не более 6 - 8 Гц мышца, состоящая из медленных двигательных единиц, отвечает одиночными сокращениями. Сокращение наступает не сразу после нанесения раздражения, а через определенный промежуток времени, называемый латентным периодом. Его величина составляет для икроножной мышцы лягушки 0,01 с. Фаза укорочения длится 0,04 с, фаза расслабления - 0,05 с.
Начало сокращения соответствует восходящей фазе потенциала действия, когда он достигает пороговой величины (примерно 40 мВ). У млекопитающих одиночное сокращение скелетных мышц длится 0,04 - 0,1 с, но оно неодинаково в различных мышцах у одного и того же животного. В красных волокнах мышц оно значительно больше, чем в белых. Если на мышцу действуют два быстро следующих друг за другом раздражения (период между импульсами не более 100 мс), мышечные волокна расслабляются не полностью и каждое последующее сокращение как бы наслаивается на предыдущее. Происходит суммация сокращений, которая может быть полной, когда оба сокращения сливаются, образуя одну вершину, или неполной, в зависимости от частоты раздражений. В обоих случаях сокращение имеет большую амплитуду, чем максимальное сокращение при одиночном раздражении.
При воздействии на мышцу ритмических раздражений высокой частоты наступает сильное и длительное сокращение мышцы, которое называется тетаническим сокращением или тетанусом. Этот термин впервые применил Э. Вебер в 1821 году.
Тетанус может быть зубчатым (при частоте раздражений 20 - 40 Гц) или сплошным, гладким (при частоте 50 Гц и выше). Амплитуда тетанического сокращения в 2 - 4 раза выше амплитуды одиночного сокращения при той же силе раздражения.
Гладкий тетанус возникает тогда, когда очередной импульс раздражения действует на мышцу до начала фазы расслабления. При очень большой частоте раздражений каждое очередное раздражение будет попадать на фазу абсолютной рефрактерности и мышца вообще не будет сокращаться. Высота мышечного сокращения при тетанусе зависит от ритма раздражения, а также от возбудимости и лабильности, которые изменяются в процессе сокращения мышцы. Тетанус наиболее высокий при оптимальном ритме, когда каждый последующий импульс действует на мышцу в фазу экзальтации, вызванной предыдущим импульсом. В этом случае создаются наилучшие условия (оптимум силы и частоты раздражения, оптимум ритма) для работы мышцы.
При тетанических сокращениях мышечные волокна утомляются больше, чем при одиночных сокращениях. Поэтому даже в пределах одной мышцы происходит периодическая смена частоты импульсации (вплоть до полного исчезновения) в разных двигательных единицах.
Импульсы с мотонейронов в условиях покоя участвуют в поддержании мышечного тонуса.
Под тонусом понимают состояние естественного постоянного напряжения мышц при невысоких энергетических затратах. В поддержании тонуса участвуют проприорецепторы мышц (мышечные веретена) и центральная нервная система.
Осуществление тонуса скелетных мышц обусловлено функцией медленных двигательных единиц красных волокон мышц. Тонус скелетных мышц связан с поступлением редких нервных импульсов к мышце, в результате чего мышечные волокна возбуждаются не одновременно, а попеременно. У домашних животных существуют специализированные рефлекторные дуги, одни из которых обеспечивают тетанические сокращения, а другие мышечный тонус. Тонус скелетных мышц играет важную роль в поддержании определенного положения тела в пространстве и деятельности двигательного аппарата.
При сближении актиновых и миозиновых фибрилл вследствие замыкания поперечных мостиков в мышечном волокне развивается напряжение (активная механическая тяга). В зависимости от условий, в которых происходит сокращение мышц, развивающееся напряжение реализуется по-разному. Различают два основных типа мышечных сокращений - изотонический и изометрический. Когда мышца при раздражении сокращается, не поднимая никакого груза, происходит укорочение мышечных волокон, но их напряжение не меняется и равно нулю, такое сокращение называют изотоническим (греч. isos - равный, tonos - напряжение). В эксперименте изотоническое сокращение получают при электрическом (тетаническом) раздражении изолированной мышцы, отягащенной небольшим грузом. Укорочение мышцы происходит при постоянном напряжении, равном внешней нагрузки.
Изометрическое (греч. isos - равный, meros - мера) - это сокращение, при котором длина волокон не уменьшается, но их напряжение возрастает (сокращение при неизменной длине). В этом случае сократительный компонент укорачивается за счет растяжения пассивного упругого компонента, который может увеличивать свою длину на 2 - 6 % от длины покоя.
С молекулярной точки зрения напряжение при изотоническом сокращении обеспечивается замыканием и размыканием поперечных мостиков. При этом скорость сокращения зависит от числа замкнутых мостиков, образуемых в единицу времени (чем их меньше, тем больше скорость и соответственно меньше сила сокращения).
При изометрическом же сокращении напряжение в мышечных волокнах создается за счет повторного прикрепления поперечных мостиков на одних и тех же фиксированных участках актиновых нитей.
В естественных условиях деятельности мышц практически не встречается чисто изотоническое или чисто изометрическое сокращение.
Смешанный тип сокращения мышц, при котором изменяются длина и напряжение, называется ауксотоническим. При совершении животным сложных двигательных актов все работающие мышцы сокращаются ауксотонически - с преобладанием либо изотонического, либо изометрического типа сокращения.
Основным элементом скелетной мышцы является мышечная клетка. В связи с тем, что мышечная клетка по отношению к своему поперечному сечению (0,05-0,11мм) относительно длинна (волокна бицепса, например, имеют длину до 15 см), ее называют также мышечным волокном.
Скелетная мышца состоит из большого количества этих структурных элементов, составляющих 85-90% от ее общей массы. Так, например, в состав бицепса входит более одного миллиона волокон.
Между мышечными волокнами расположена тонкая сеть мелких кровеносных сосудов (капилляров) и нервов (приблизительно 10% от общей массы мышцы). От 10 до 50 мышечных волокон соединяются в пучок. Пучки мышечных волокон и образуют скелетную мышцу. Мышечные волокна, пучки мышечных волокон и мышцы окутаны соединительной тканью.
Мышечные волокна на своих концах переходят в сухожилия. Через сухожилия, прикрепленные к костям, мышечная сила воздействует на кости скелета. Сухожилия и другие эластичные элементы мышцы обладают, кроме того, и упругими свойствами. При высокой и резкой внутренней нагрузке (сила мышечной тяги) или при сильном и внезапном внешнем силовом воздействии эластичные элементы мышцы растягиваются и тем самым смягчают силовые воздействия, распределяя их в течение более продолжительного промежутка времени.
Поэтому после хорошей разминки в мускулатуре редко происходят разрывы мышечных волокон и отрывы от костей. Сухожилия обладают значительно большим пределом прочности на растяжение (около 7000 Н/кв см), чем мышечная ткань (около 60 Н/кв см), где Н – ньютон, поэтому они гораздо тоньше, чем брюшко мышцы. В мышечном волокне содержится основное вещество, называемое саркоплазмой. В саркоплазме находятся митохондрии (30-35% от массы волокна), в которых протекают процессы обмена веществ и накапливаются вещества, богатые энергией, например фосфаты, гликоген и жиры. В саркоплазму погружены тонкие мышечные нити (миофибриллы), лежащие параллельно длинной оси мышечного волокна.
Миофибриллы составляют в совокупности приблизительно 50% массы волокна, их длина равна длине мышечных волокон, и они являются, собственно говоря, сократительными элементами мышцы. Они состоят из небольших, последовательно включаемых элементарных блоков, именуемых саркомерами (рис. 33).
Рис. 33. Схема скелетной мышцы: мышца (до 5 см), пучок мышечных волокон (0,5 мм), мышечное волокно (0,05-0,1 мм), миофибрилла (0,001-0,003 мм). Цифры в скобках обозначают приблизительный размер поперечного сечения строительных элементов мышцы
Так как длина саркомера в состоянии покоя равна приблизительно лишь 0,0002 мм, то для того, чтобы, к примеру, образовать цепочки из звеньев миофибрилл бицепса длиной 10-15 см, необходимо "соединить" огромное количество саркомеров. Толщина мышечных волокон зависит главным образом от количества и поперечного сечения миофибрилл.
В миофибриллах скелетных мышц наблюдается правильное чередование более светлых и более темных участков. Поэтому часто скелетные мышцы называют поперечнополосатыми. Миофибрилла состоит из одинаковых повторяющихся элементов, так называемых саркомеров. Саркомер ограничен с двух сторон Z-дисками. К этим дискам с обеих сторон прикрепляются тонкие актиновые нити. Нити актина обладают низкой плотностью и поэтому под микроскопом кажутся более прозрачными или более светлыми. Эти прозрачные, светлые области, располагающиеся с обеих сторон от Z-диска, получили название изотропных зон (или I-зон).
В середине саркомера располагается система толстых нитей, построенных преимущественно из другого сократительного белка, миозина. Эта часть саркомера обладает большей плотностью и образует более темную анизотропную зону (или А-зону). В ходе сокращения миозин становится способным взаимодействовать с актином и начинает тянуть нити актина к центру саркомера. Вследствие такого движения уменьшается длина каждого саркомера и всей мышцы в целом. Важно отметить, что при такой системе генерации движения, получившей название системы скользящих нитей, не изменяется длина нитей (ни нитей актина, ни нитей миозина). Укорочение является следствием лишь перемещения нитей друг относительно друга. Сигналом для начала мышечного сокращения является повышение концентрации Са 2+ внутри клетки. Концентрация кальция в клетке регулируется с помощью специальных кальциевых насосов, встроенных в наружную мембрану и мембраны саркоплазматического ретикулума, который оплетает миофибриллы.
Двигательная единица (ДЕ) – группа мышечных волокон, иннервируемых одним мотонейроном. Мышца и ее нервный привод состоят из большого количества параллельно расположенных ДЕ (рис. 34).

Рис. 34. Строение двигательной единицы: 1 – спинной мозг; 2 – мотонейроны; 3 – аксоны; 4 – мышечные волокна
В нормальных условиях ДЕ работает как единое целое: посылаемые мотонейроном импульсы приводят в действие все входящие в ее состав мышечные волокна. Благодаря тому, что мышца состоит из множества ДЕ (в крупных мышцах до несколько сотен), она может работать не всей массой, а по частям. Это свойство используется при регуляции силы и скорости мышечного сокращения. В естественных условиях частота импульсов, посылаемых мотонейронами в ДЕ, находится в пределах 5–35 имп./с, лишь при максимальных мышечных усилиях удается зарегистрировать частоту разрядов выше 50 имп./с.
Компоненты ДЕ обладают различной лабильностью: аксон – до 1000 имп./с, мышечное волокно – 250-500, мионевральный синапс – 100–150, тело мотонейрона – до 50 имп./с. Утомляемость компонента тем выше, чем меньше его лабильность.
Различают быстрые и медленные ДЕ. Быстрые обладают большой силой и скоростью сок-ращения в короткое время, высокой активностью гликолитических процессов, медленные рабо-тают в условиях высокой активности окислительных процессов длительно, при меньшей силе и скорости сокращения. Первые быстро утомляемы, содержат много гликогена, вторые выносливы – в них много митохондрий. Медленные ДЕ активны при любом напряжении мышцы, тогда как быстрые ДЕ активны лишь при сильных мышечных напряжениях.
Основываясь на анализе ферментов мышечных волокон, их классифицируют на три вида: тип I, тип IIа, тип IIб.
В зависимости от скорости сокращения, аэробной и анаэробной возможности используют понятия: медленно-сокращающийся, окислительный тип (МО), быстро-сокращающийся, окислительно-гликолитический тип (БОГ) и быстро-сокращающийся, гликолитический тип (БГ).
Существуют и другие классификации ДЕ. Так, основываясь на двух параметрах – снижении прерывистого тетануса и сопротивлении утомлению – ДЕ делят на три группы (Burke, 1981): медленно сокращающиеся, невосприимчивые к утомлению (тип S); быстро сокращающиеся невосприимчивые к утомлению (тип FR) и быстро сокращающиеся восприимчивые к утомлению (тип FF).
Волокна I типа соответствуют волокнам типа МО, волокна IIа типа– волокнам типа БОГ, а волокна IIб типа– волокнам типа БГ. Мышечные волокна типа МО относятся к ДЕ типа S, волокна типа БОГ – к ДЕ типа FR, а волокна типа БГ – к ДЕ типа FF.
Каждая мышца человека содержит совокупность всех трех типов волокон. ДЕ типа FF характеризуется наибольшей силой сокращения, наименьшей продолжительностью сокращения и наибольшей восприимчивостью к утомлению.
Говоря о пропорциях различных мышечных волокон у человека, следует отметить, что и у мужчин, и у женщин несколько больше медленных
волокон (по данным различных авторов –
от 52 до 55%).
Имеется строгая зависимость между количеством медленно- и быстро сокращающихся волокон в мышечной ткани и спортивными достижениями на спринтерских и стайерских дистанциях.
Икроножные мышцы чемпионов мира по марафону содержат 93–99% медленных волокон, тогда как у сильнейших спринтеров мира в этих мышцах больше количество быстрых волокон (92%).
У нетренированного человека число двигательных единиц, которые могут быть мобили-зованы при максимальных силовых напряжениях, обычно не превышает 25–30%, а у хорошо тренированных к силовым нагрузкам лиц число вовлеченных в работу моторных единиц может превышать 80–90%. В основе этого явления лежит адаптация центральной нервной системы, приводящая к повышению способности моторных центров мобилизовывать большее число мотонейронов и к совершенствованию межмышечной координации (рис. 35).

Рис. 35. Характеристика двигательных единиц
Физиология двигательного аппарата.
ЛЕКЦИЯ № 15
Потребность организма в кислороде
При многих состояниях, включая упомянутые выше, кислород назначается в лечебных целях. В тех случаях, когда поступление О 2 прекращается более чем на 4 минуты, в мозге происходят необратимые изменения и человек умирает. Подобная ситуация возникает, например, когда ребенок, играя с полиэтиленовым пакетом, надевает его себе на голову и задыхается. Если поступление СО 2 только уменьшается, может развиваться гипоксия мозга . Обычно это происходит с людьми, работающими в ограниченном пространстве (трюмах, цистернах, котлах). В этих условиях они быстро используют имеющийся в распоряжении воздух и могут погибнуть от аноксии , если их дополнительно не обеспечить кислородом или не вывести на свежий воздух.
При недостатке кислорода кровь теряет присущий ей ярко красный цвет и приобретает голубоватый оттенок. Одновременно губы пациента, ушные раковины и конечности становятся цианотическими , то есть синюшного цвета.
У человека существует три вида мышц (Рис. 32):
Ø поперечно-полосатые скелетные мышцы составляют 30-35% массы тела и имеют площадь около 3 м 2 . Целая мышца – отдельный орган, а мышечное волокно – отдельная клетка (Рис. 33);
Ø особая поперечно-полосатая сердечная мышца;
Ø гладкие мышцы внутренних органов.
Рис. 32. Виды мышечной ткани: I- продольный разрез; II- поперечный срез; А - гладкая (неисчерченная); Б - поперечнополосатая скелетная; В - поперечнополосатая сердечная
Мышцы иннервируются тремя способами:
Ø двигательными нервами, которые передают из центра моторные команды;
Ø чувствительными нервами, информация по которым передается в центр о напряжении и движении мышц;
Ø симпатическими нервными волокнами, которые влияют на обменные процессы в мышцах.
Функции скелетных мышц:
– перемещение частей тела друг относительно друга, фиксация внутренностей;
– перемещение тела в пространстве (локомоция);
– поддержание позы;
– участите в обмене веществ, терморегуляции и поддержания тонуса нервной и сердечно-сосудистой систем.

Рис. 33. Схема скелетной мышцы: А - мышечные волокна прикреплены к сухожилиям; Б- отдельное волокно, состоящее из миофибрилл; В- отдельная миофибрилла: чередование светлых актиновых I-дисков и темных миозиновых А-дисков; наличие Н-зоны и М-линии; Г- поперечные мостики между толстыми миозиновыми и тонкими актиновыми нитями
Функциональной единицей скелетной мышцы является двигательная единица, которая состоит из мотонейрона спинного мозга, его аксона (двигательного нерва) с многочисленными окончаниями, и иннервируемых им мышечных волокон. Возбуждение мотонейрона вызывает одновременное сокращение всех входящих в эту единицу мышечных волокон. Двигательные единицы (ДЕ) небольших мышц содержат мало мышечных волокон (ДЕ глазного яблока 3-6 волокон), ДЕ крупных мышц туловища и конечностей – около 2000 волокон.
Мышечное волокно представляет собой вытянутую клетку длиной 10-12 см (длина мышечного волокна обычно равна длине самой мышцы), диаметр волокна около 10-100 мкм. В состав мышечного волокна входит (Рис. 33):
· Оболочка – сарколемма.
· Жидкое содержимое – саркоплазма.
· Митохондрии – энергетические центры клетки.
· Рибосомы – белковые депо.
· Миофибриллы (фибриллы) – сократительные элементы, кторые состоят из 2 видов белков (тонкие нити актина и вдвое более толстые нити миозина). Миофибриллы разделены Z – мембранами (или Z – линиями) на отдельные участки – саркомеры, в средней части которых расположены преимущественно миозиновые нити (толстые филаменты), а актиновые нити (тонкие филаменты) прикреплены к Z – мембранам по бокам саркомера (разная способность преломлять свет у актина и миозина создает в состоянии покоя мышцы поперечно-полосатый вид в световом микроскопе). Темные участки называются А-диски, светлые I-диски. В средней части А-диска есть более светлый участок – Н-зона. В покоящейся мышце в Н-зоне нет тонких филаментов, а в I-диске нет толстых.
· Саркоплазматический ретикулум – замкнутая система продольных трубочек и цистерн, расположенных вдоль миофибрилл и содержащих ионы Са 2+
Толстые филаменты состоят примерно из 400 молекул миозина (закрученных друг относительно друга), который имеет вид палочковидной молекулы с утолщенным концом – головкой (Рис. 33, Г).
Тонкие филаменты образованы тремя белками (Рис. 34):
- актин – глобулярный белок, который образует спиральный двухнитевой полимер, состоящий из 13-14 молекул;
- тропомиозин – палочковидная молекула, которая располагается в желобке двойной спирали актина, длина молекулы тропомиозина равна длине 7 мономеров актина
- тропонин – сферическая молекула состоит из 3 субъединиц (ТнС, ТнТ, ТнI): Са-связывающая, тропомиозинсвязывающая и ингибирующая.