Selgroogsete vereringesüsteemid (kompleks). Kalade vereringesüsteem Kalade vereringesüsteem mitu vereringeringi
Selle ja kehakudede vahel toimub ainete vahetus. Veresoonte säng on väga pikk ja sellel on palju harusid, mis häirivad normaalset verevoolu. See tähendab, et kogu tee läbimiseks on vaja kehtestada teatud surve ja see on see, mida süda loob.
Selle organi ehitus kaladel on lihtsam kui maismaaloomadel. Teades, kui palju on kaladel ja kui palju teistel olenditel, on võimalik teha võrdlev analüüs. See võimaldab teil selgelt näha nende kardiovaskulaarsüsteemi erinevusi ja sarnasusi.
Mitu südamekambrit on kaladel?
Kalade süda on väikese kaaluga, vaid 0,1% nende kehakaalust, kuigi sellest reeglist on ka erandeid. Ja paljud inimesed mäletavad oma kooliajast, kui palju südamekambreid on kaladel. Neid on ainult kaks - aatrium ja vatsake. Kuid neil on struktuuris erinevusi. Üldise skeemi kohaselt on kahte tüüpi sarnasusi ja erinevusi.
Mõlemal valikul on neli õõnsust:
- venoosne siinus;
- klapiga aatrium;
- vatsakese;
- teatud moodustis, mille struktuur sarnaneb aordikaarega.
Elasmobranchil on arteriooskoonus, teleostidel aga bulbus arteriosus. Nende skeemide erinevus seisneb arteriaalsete moodustiste ja vatsakeste morfofunktsionaalsetes omadustes. Esimesel juhul on kaladel kiuline kude ilma klappideta. Elasmobranchide hulka kuuluvatel kaladel sisaldab arteriooskoonus lihaskudet ja klapisüsteemi.

Seda kõike teades saavad kõik teada, kui palju südamekambreid on kaladel ja milline on nende struktuur. Eriti huvitav on müokardi struktuur, kuna seda esindab homogeenne südamekude. See on teistest loomadest õhem.
Südame töö
Selle järgi, mitu kambrit on kala südamel, saab määrata selle organi tööpõhimõtte ja selle rütmid. Südame löögisageduse (HR) määravad paljud tegurid, sealhulgas vee temperatuur ja kala vanus.
Selguse huvides tehakse ettepanek võtta arvesse karpkala pulssi toatemperatuuril vees.
Teadlased jõudsid järeldusele, et kokkutõmmete sagedust mõjutab tugevalt vee temperatuur. Mida külmem on tiik, seda aeglasemalt lööb süda. Niisiis, temperatuuril 8 °C on pulss ligikaudu 25 lööki minutis ja temperatuuril 12 °C - 40 lööki.
Tiraaž
Teades, milline süda on kalal ja mitu kambrit selles on, võib ette kujutada, kui palju neil vereringeringe on. Tänu sellele, et seal on kaks kambrit, on kaladel ainult üks vereringe ring, kuigi veri ringleb selle kaudu pikka aega. Täisringi läbimiseks kulub umbes kaks minutit ja inimestel teeb veri kaks ringi 23 sekundiga.
Vere liikumine algab vatsakesest. Sealt läheb see läbi pirni või arterioosse koonuse kõhuaordi. Veri jaguneb kaheks kanaliks, mis ulatuvad lõpuseniitideni. Kroonlehearterist väljuvad kaks arteriooli, mis moodustavad kapillaaride võrgustiku. See ühineb üheks eferentseks arteriooliks, mis läheb eferentsesse kroonlehearterisse. Viimased moodustavad parema ja vasaku eferentse haruarteri.
Unearterid ulatuvad peani ja lõpusearterid moodustavad seljaaordi, mis kulgeb mööda kogu kala selgroolüli. Läbinud kogu keha, naaseb veri venoosse voodi kaudu südamesse venoossesse siinusesse. Kala südame struktuur võimaldab pumbata ainult venoosset verd. Lõpuseaparaati läbides vahetab venoosne veri gaase veega.
Kalade vereringesüsteemi anumatel on klapiaparaat. See takistab vere tagasivoolu läbi kanali. Selle liikumise ühtsuse tagab südame ühtlane täitumine, ilma järskude kõikumisteta, mida inimestel täheldatakse.

Lõpuks
Struktuur on lihtne. Sellel on ainult kaks kambrit: aatrium ja vatsake. Elundi ühtlane täitumine verega ja veresoonte tugev hargnemine pikendavad aega, mis kulub vere ringlemiseks. Veelgi enam, külmas vees kulub vere ringlemiseks kauem aega.
Kahepaiksetel toimub seoses põhimõtteliselt uue elupaiga kujunemise ja osalise üleminekuga õhuhingamisele vereringesüsteem mitmeid olulisi morfofüsioloogilisi muutusi: neil on teine vereringe ring.
Konna süda asub keha esiosas, rinnaku all. See koosneb kolmest kambrist: vatsakesest ja kahest kodadest. Nii kodade kui ka seejärel vatsakese tõmbuvad kokku vaheldumisi.

Kuidas konna süda töötab?
Vasak aatrium saab kopsudest hapnikuga rikastatud arteriaalse verd ja parem aatrium süsteemsest vereringest venoosset verd. Kuigi vatsakest ei ole jagatud, ei segune need kaks verevoolu peaaegu üldse (vatsakese seinte lihaste väljakasvud moodustavad üksteisega suhtlevaid kambreid, mis takistavad vere täielikku segunemist).
Vatsake erineb teistest südameosadest paksude seinte poolest. Selle sisepinnalt ulatuvad pikad lihasepaelad, mis on kinnitatud kahe atrioventrikulaarset (atrioventrikulaarset) ava katva klapi vabade servade külge, mis on ühised mõlemale kodale. Arteriooskoonus on varustatud klappidega põhjas ja otsas, kuid lisaks on selle sees pikk pikisuunaline spiraalklapp.
Vatsakese paremalt küljelt väljub arteriaalne koonus, mis jaguneb kolmeks arterikaare paariks (kopsu naha-, aordi- ja unekaared), millest igaüks väljub sellest iseseisva avaga. Vatsakese kokkutõmbumisel surutakse esmalt välja kõige vähem oksüdeerunud veri, mis läbi naha kopsukaarte siseneb gaasivahetuseks (kopsuringluseks) kopsudesse. Lisaks saadavad kopsuarterid oma oksad nahka, mis samuti osaleb aktiivselt gaasivahetuses. Järgmine osa segaverest saadetakse süsteemsetesse aordikaaredesse ja edasi kõikidesse keha organitesse. Enim hapnikuga küllastunud veri voolab unearteritesse, mis varustavad aju. Conus arteriosuse spiraalklapp mängib sabata kahepaiksete verevoolu eraldamisel suurt rolli.
Konna puhul voolab veri südame vatsakesest arterite kaudu kõikidesse organitesse ja kudedesse ning neist veenide kaudu paremasse aatriumisse - see on suur vereringe ring.
Lisaks voolab veri vatsakesest kopsudesse ja nahka ning kopsudest tagasi südame vasakusse aatriumisse - see on kopsuvereringe. Kõigil selgroogsetel, välja arvatud kaladel, on kaks vereringeringi: väikesed - südamest hingamisorganitesse ja tagasi südamesse; suur - südamest arterite kaudu kõikidesse organitesse ja neist tagasi südamesse.
Sarnaselt teistele selgroogsetele imbub kahepaiksete vere vedel fraktsioon läbi kapillaaride seinte rakkudevahelisse ruumi, moodustades lümfi. Konnade naha all on suured lümfikotikesed. Neis tagavad lümfi voolu spetsiaalsed struktuurid, nn. "lümfisüdamed". Lõpuks kogutakse lümf lümfisoontesse ja suunatakse tagasi veenidesse.
Seega, kuigi kahepaiksetel moodustub kaks vereringeringi, ei ole need tänu ühele vatsakesele täielikult eraldatud. See vereringesüsteemi struktuur on seotud hingamiselundite duaalsusega ja vastab selle klassi esindajate kahepaiksete elustiilile, võimaldades viibida maal ja veeta pikka aega vees.
Kahepaiksete vastsetel on üks vereringesüsteem (sarnane kalade vereringesüsteemiga). Kahepaiksetel ilmub uus vereloomeorgan - torukujuliste luude punane luuüdi. Nende vere hapnikumaht on suurem kui kaladel. Kahepaiksete punased verelibled on tuumalised, kuid neid on vähe, kuigi need on üsna suured.

Kahepaiksete, roomajate ja imetajate vereringesüsteemi erinevused
Kahepaiksete hingamissüsteem mida esindavad kopsud ja nahk, mille kaudu nad saavad ka hingata. Kopsud- need on paaris õõnsad kotid, mille rakuline sisepind on täpiline kapillaaridega. Siin toimub gaasivahetus. Konnade hingamismehhanism viitab süstimisele ja seda ei saa nimetada täiuslikuks. Konn tõmbab õhku orofarüngeaalsesse õõnsusse, mis saavutatakse suupõhja langetamise ja ninasõõrmete avamisega. Seejärel tõuseb suupõhi ja ninasõõrmed sulguvad uuesti klappidega ning õhk surutakse kopsudesse.
Konna vereringesüsteem sisaldab kolmekambriline süda(kaks kodat ja vatsake) ja kaks ringlus- väike (kopsu) ja suur (pagasiruumi). Kahepaiksete kopsuvereringe algab vatsakesest, läbib kopsuveresooned ja lõpeb vasakpoolses aatriumis.

Süsteemne vereringe See algab ka vatsakesest, läbib kõik kahepaikse keha veresooned ja naaseb paremasse aatriumi. Nagu imetajatelgi, on veri kopsudes hapnikuga küllastunud ja seejärel kannab seda kogu kehas.
Vasak aatrium saab arteriaalset verd kopsudest ja parempoolne venoosne veri ülejäänud kehast. Samuti siseneb veri paremasse aatriumisse, mis läbib naha pinna ja on seal hapnikuga küllastunud.
Hoolimata asjaolust, et nii venoosne kui arteriaalne veri siseneb vatsakesse, ei segune see seal klappide ja taskute süsteemi olemasolu tõttu täielikult. Selle tõttu läheb arteriaalne veri ajju, venoosne veri nahka ja kopsudesse ning segaveri teistesse organitesse. Just segavere olemasolu tõttu on kahepaiksete eluprotsesside intensiivsus madal ja kehatemperatuur võib sageli muutuda.
Lisamaterjalid teemal: Kahepaiksete hingamis- ja vereringesüsteem.
Maojalgsete klass
Kahepaiksed (Amphibians) klass.
Kahepaiksetel on väike ringlus
 |
Konna arteriaalse süsteemi skeem (hõreda varjundiga on kujutatud rohkem arteriaalset verd, tihedama varjundiga segaverd, musta värviga venoosset verd):
1 – parem aatrium,
2 - vasak aatrium,
3 - vatsake,
4 – arteriaalne koonus,
5 - naha-kopsu
6 - kopsuarter,
7 - nahaarter,
8 - parem aordikaar,
9 - vasak aordikaar,
10 - kuklaluu arter, 11 - subklaviaarter, 12 - dorsaalne aort, 13 - enteromesenteerne arter,
14 - urogenitaalarterid, 15 - ühine niudearter,
16 - ühine unearter, 17 - sisemine unearter,
18 - väline unearter, 19 - kops, 20 - maks,
21 - magu, 22 - sooled, 23 - munandid, 24 - neer
Konna venoosse süsteemi skeem(rohkem arteriaalset verd on näha hõreda varjundiga, segaverd täppidega, venoosset verd musta värviga):
1 - venoosne siinus,
2 - parem aatrium,
3 - vasak aatrium,
4 - vatsake,
5 - reieluu veen,
6 - istmikunärvi veen,
7 - neerude portaalveen,
8 - kõhu veen,
9 - maksa portaalveen, 10 - eferentne neeruveen
11 - tagumine õõnesveen, 12 - maksa veen,
13 - suur nahaveen, 14 - õlavarre veen,
15 - subklavia veen, 16 - välimine kägiveen,
17 - sisemine kägiveen, 18 - parem eesmine õõnesveen, 19 - vasak eesmine õõnesveen, 20 - kopsuveenid, 21 - kops, 22 - maks, 23 - neer, 24 - munandid,
25 - magu, 26 - sooled
Kas te ei leidnud seda, mida otsisite? Kasutage otsingut:
Loe ka:
Konna siseehituse uurimine
Märgpreparaadil uurige siseorganite asukohta (joonis 21). Leidke see keha rindkere piirkonnast süda. Leidke koda ja vatsake: kodade värvus on tumedam, vatsake on hele, selle seinad on lihaselisemad (joonis 22).
Tutvuge suurema ja väiksema ringlusega diagrammiga (joonis 23). Südamest paremal ja vasakul asuvad kopsud. Kui kopsud on õhuga täidetud, näevad need välja nagu suured helehallid kotid. Konna hingamismehhanism on survetüüpi (joon. 24).
Otsi suguelundid emased - munasarjad, munajuhad. Munajuhad on pikad värvilised torud. Meestel munandid kollakasvalge, oakujuline. Iga munand on ühendatud neeru ja kusejuhaga, seetõttu toimivad konna kusejuhad ka vasdeferenidena (Wolffi kanal).

Riis. 21. Emaskonna siseorganite üldine paigutus:
1 - parem aatrium, 2 - vasak aatrium, 3 - vatsakese, 4 - arteriaalne koonus, 5 - valgus, 6 - söögitoru, 7 - kõht, 8 - mao pülooriline osa, 9 - kaksteistsõrmiksool, 10 - pankreas, 11 - peensoolde, 12 - pärasool, 13 - kloaagi piirkond, 14 - maks, 15 - sapipõis, 16 - sapijuha, 17 - mesenteeria, 18 - põrn, 19 - neerud, 20 - kusejuha, 21 - põis, 22 - munasarjad, 23 – munajuha (vasakut munasarja ja munajuha pole joonisel kujutatud).
 Riis. 22.
Riis. 22.
Kahepaiksete kopsuvereringe lõpeb
Avatud konnasüdame skeem:
1 - parem aatrium, 2 - vasak aatrium, 3 - vatsakese, 4 – klapid, mis sulgevad mõlemast kodadest vatsakesse viiva ühise ava, 5 - arteriaalne koonus, 6 - ühine arteritüvi, 7 - naha kopsuarter, 8 - aordi kaar, 9 - ühine unearter, 10 - karotiidnääre 11 – arteriooskoonuse spiraalklapp.

Riis. 23. Vereringe kahepaiksetel:
 A– kulles (ühe tsirkulatsiooniga vastne), B- täiskasvanu (kahe vereringeringiga), I, II, III, IV- haruarterite arteriaalsed kaared, 1
- parem aatrium, 2
- vasak aatrium, 3
- vatsakese, 4
- arteriaalne koonus, 5
- aordi juured, 6
- dorsaalne aort, 7
- lõpused, 8
- unearterid, 9
- kopsud, 10
- veenid, mis toovad arteriaalset verd kopsudest, 11
- kopsuarterid, mis kannavad venoosset verd südamest, 12
- veenid, mis toovad veeniverd kogu kehast, 13
– sulanud II ja III arterikaared, mis kannavad südamest segaverd. Venoosne veri on näidatud musta värviga, arteriaalne veri on valge, segaveri on teritatud.
A– kulles (ühe tsirkulatsiooniga vastne), B- täiskasvanu (kahe vereringeringiga), I, II, III, IV- haruarterite arteriaalsed kaared, 1
- parem aatrium, 2
- vasak aatrium, 3
- vatsakese, 4
- arteriaalne koonus, 5
- aordi juured, 6
- dorsaalne aort, 7
- lõpused, 8
- unearterid, 9
- kopsud, 10
- veenid, mis toovad arteriaalset verd kopsudest, 11
- kopsuarterid, mis kannavad venoosset verd südamest, 12
- veenid, mis toovad veeniverd kogu kehast, 13
– sulanud II ja III arterikaared, mis kannavad südamest segaverd. Venoosne veri on näidatud musta värviga, arteriaalne veri on valge, segaveri on teritatud.

Riis. 24. Konna hingamismehhanismi skeem:
I- suuõõs laieneb ja õhk siseneb sellesse avatud ninasõõrmete kaudu, II- ninasõõrmed sulguvad, kõripilu avaneb ja kopsudest väljuv õhk seguneb suuõõnes atmosfääriõhuga, III- ninasõõrmed on suletud, suuõõs tõmbub kokku ja segaõhk surutakse kopsudesse, IV– kõrilõhe suletakse, suuõõne põhi surutakse vastu suulae, surudes ülejäänud õhu läbi avatud ninasõõrmete välja: 1 - ninasõõrme välimine avamine, 2 - ninasõõrme sisemine avamine (choana), 3 - suuõõne, 4 - suuõõne põrand, 5 - kõrilõhe, 6 - valgus, 7 - söögitoru.
Riis. 25. Emaskonna kloaagi skeem: 1 - kloaagi välimine avamine, 2 - kloaagi õõnsus, 3 - pärasool, 4 - põis, 5 - kusejuha, 6 - munajuha, 7 - vaagna sein.
 Laiendatud kõht kaetud maksa vasaku sagaraga. See algab temast kaksteistsõrmiksool. Tema loop on kõhunääre. Kaksteistsõrmiksool muutub järk-järgult õhuke, moodustades mitu silmust, jätkub viimane sisse rasv. Soolestik lõpeb kloaak(Joonis 25). Soolestiku uurimisel ärge ajage neid segamini munajuhade aasadega.
Laiendatud kõht kaetud maksa vasaku sagaraga. See algab temast kaksteistsõrmiksool. Tema loop on kõhunääre. Kaksteistsõrmiksool muutub järk-järgult õhuke, moodustades mitu silmust, jätkub viimane sisse rasv. Soolestik lõpeb kloaak(Joonis 25). Soolestiku uurimisel ärge ajage neid segamini munajuhade aasadega.
Suguküpsel emasel silmatorkav munasarjad– suured tumedat värvi kärgkotid. Vasakul oleva munasarja all on näha lülisamba külgedel neerud- tumepunase värvi spindlikujulised moodustised. Kuidas kalad seda teevad? mesonefros.
Nad liiguvad neist eemale kusejuhad, voolab sisse kloaak, ja põis avaneb kloaaki eraldi avaga (joon. 26 ja 27).
Munandite ja munasarjade ülemises osas on erekollase või oranži värvi lobed moodustised. Need on rasvakehad, mis sisaldavad palju toitaineid, mis on vajalikud paljunemisproduktide arendamiseks.
Riis. 26. Emase konna urogenitaalsüsteem:
 1
- neerud, 2
- kusejuha, 3
- kloaagi õõnsus, 4
- kuseteede avamine, 5
- põis, 6
- põie avamine, 7
- vasak munasari (paremat munasarja pole joonisel näidatud), 8
- munajuha, 9
- munajuha lehter, 10
- rasvane keha (parema külje rasvast keha pole näidatud), 11
- neerupealised, 12
– suguelundite avamine (munajuha avamine).
1
- neerud, 2
- kusejuha, 3
- kloaagi õõnsus, 4
- kuseteede avamine, 5
- põis, 6
- põie avamine, 7
- vasak munasari (paremat munasarja pole joonisel näidatud), 8
- munajuha, 9
- munajuha lehter, 10
- rasvane keha (parema külje rasvast keha pole näidatud), 11
- neerupealised, 12
– suguelundite avamine (munajuha avamine).
Riis. 27. Isase konna urogenitaalsüsteem:
1 - neerud, 2 - kusejuha (tuntud ka kui vas deferens), 3 - kloaagi õõnsus, 4 - urogenitaalne avamine, 5 – põis, 6 – põie avamine, 7 - munandid, 8 - seemnetorukesed, 9 - seemnepõiekesed, 10 - rasvane keha, 11 - neerupealised.

Riis. 28. Konnaaju ülalt ( A) ja allpool ( B)
1 - eesaju ajupoolkerad, 2 - lõhnasagar, 3 - haistmisnärv 4 - vahepea, 5 - visuaalne chiasma, 6 - lehter, 7 - ajuripats, 8 - keskaju optilised sagarad, 9 - väikeaju, 10 - medulla, 11 - selgroog.

Riis. 29. Konna luustik:
I- terve skelett, II- selgroolüli ülal, III- selgroolüli ees, 1 - kaelalüli, 2 - ristluu lüli, 3 - urostiil, 4 - rinnaku, 5 - kõhreline rinnaku tagumine osa, 6 - presternum, 7 - korakoid, 8 - prokorakoid, 9 - abaluu, 10 - abaluuülene kõhr, 11 - ilium, 12 - ischium, 13 - häbemekõhre, 14 - õlavarreluu, 15 - küünarvars (raadius + küünarluu), 16 - ranne, 17 - kämblaluu, 18 - algeline esimene sõrm, 19 - II sõrm, 20 - V sõrm, 21 - puus, 22 - sääreluu (sääreluu ja pindluu), 23 - tarsus, 24 - metatarsus, 25 - täiendava sõrme alge, 26 - ma sõrm, 27 - selgroog, 28 - seljaaju kanal, 29 - liigendatud platvorm, 30 - põikprotsess.
kesknärvisüsteem. Progresseeruvad ehituslikud tunnused: kahepaiksete eesaju on suurem kui kaladel, selle poolkerad on täielikult eraldatud (joon. 28).
Skelett Uurige preparaadil olevaid konni ja võrrelge neid pildiga (joonis 29).
Progresseeruvad märgid:
1) viiesõrmelised vabad jäsemed,
2) vööde ja jäsemete moodustamine,
3) lülisamba suurem diferentseerumine.
Primitiivsed omadused:
1) kolju kerge luustumine,
2) emakakaela ja ristluu piirkonna halb areng,
3) ribide puudumine.
Konna elupaik
Konnad elavad niisketes kohtades: soodes, märgades metsades, niitudel, mageveekogude kallastel või vees. Konnade käitumise määrab suuresti niiskus. Kuiva ilmaga varjuvad mõned konnaliigid end päikese eest, kuid pärast loojumist või märja vihmase ilmaga on neil aeg jahti pidada.
Mitu tsirkulatsiooni on kahepaiksetel?
Teised liigid elavad vees või vee lähedal, seega jahivad nad päeval.
Konnad toituvad erinevatest putukatest, peamiselt mardikatest ja kahevõsudest, kuid söövad ka ämblikke, maismaatigusid, vahel ka kalamaimusid. Konnad varitsevad oma saaki, istuvad liikumatult eraldatud kohas.
Jahipidamisel mängib suurt rolli nägemine. Märganud putukat või muud väikest looma, viskab konn suust välja laia kleepuva keele, mille külge ohver kinni jääb. Konnad haaravad ainult liikuvat saaki.

Joonis: Konna keele liikumine
Konnad on aktiivsed soojal aastaajal. Sügise algusega lahkuvad nad talveks. Näiteks talvitub rohukonn jäävabade veehoidlate põhjas, jõgede ja ojade ülemjooksul, kuhjudes kümnete ja sadade isenditega. Terava näoga konn ronib talveks mullapragudesse.
Konna väline struktuur
Konna keha on lühike, suur lame pea ilma teravate piirideta sulandub kehasse. Erinevalt kaladest on kahepaiksete pea kehaga liikuvalt liigendatud. Kuigi konnal pole kaela, võib ta pead kergelt kallutada.

Joonis: Konna välisehitus
Peas on näha kaks suurt punnis silma, kaitstud sajandite jooksul: nahkjas - pealis ja läbipaistev liigutatav - alumine. Konn pilgutab sageli silmi, samal ajal kui silmalaugude niiske nahk niisutab silmade pinda, kaitstes neid kuivamise eest. See omadus arenes konnal välja seoses tema maise eluviisiga. Kaladel, kelle silmad on pidevalt vees, silmalauge pole. Peas on silmade ees paar ninasõõret näha. Need ei ole ainult haistmisorganite avad. Konn hingab atmosfääriõhku, mis siseneb tema kehasse ninasõõrmete kaudu. Silmad ja ninasõõrmed asuvad pea ülaosas. Kui konn vette peitub, ajab ta nad välja. Samal ajal saab ta hingata atmosfääriõhku ja näha, mis toimub väljaspool vett. Konna pea iga silma taga on väike nahaga kaetud ring. See on kuulmisorgani välimine osa - kuulmekile. Konna sisekõrv, nagu kalalgi, asub kolju luudes.
Konnal on hästi arenenud paarisjäsemed – esi- ja tagajalad. Iga jäse koosneb kolmest põhiosast. Esijalas on: õlg, küünarvarre Ja pintsel. Konna käsi lõpeb nelja sõrmega (tema viies sõrm on vähearenenud). Tagajäsemes nimetatakse neid sektsioone puusa, säär, jalg. Jalg lõpeb viie varbaga, mis on konnal ühendatud ujumismembraaniga. Jäsemete osad on liigutatavalt liigendatud üksteisega kasutades liigesed. Tagajalad on palju pikemad ja tugevamad kui esijalad, neil on liikumises suur roll. Istuv konn toetub kergelt kõverdatud esijäsemetele, tagajäsemed on aga volditud ja paiknevad keha külgedel. Neid kiiresti sirgendades teeb konn hüppe. Esijalad kaitsevad looma vastu maapinda. Konn ujub tagajäsemeid tõmmates ja sirutades, surudes samal ajal esijäsemeid kehale.
Kõigil tänapäevastel kahepaiksetel on alasti nahk. Konnal on see alati niiske tänu nahanäärmete vedelatele limaskestadele.
Vesi keskkonnast (reservuaaridest, vihmast või kastest) satub konna kehasse läbi naha ja koos toiduga. Konn ei joo kunagi.
Konna luustik
Konna luustik koosneb samadest põhiosadest nagu ahvena luustik, kuid oma poolmaise eluviisi ja jalgade arengu tõttu erineb ta mitmete tunnuste poolest.

Muster: Konna luustik
Erinevalt kaladest on konnal kaelalüli. See on koljuga liikuvalt liigendatud. Sellele järgnevad külgmiste protsessidega tüvelülid (konna ribid ei ole arenenud). Kaela- ja kerelülidel on kõrgemad kaared, mis kaitsevad seljaaju. Konna ja kõigi teiste sabata kahepaiksete selgroo lõpus on pikk sabaluu. Vesilastel ja teistel sabaga kahepaiksetel koosneb see selgroo osa suurest hulgast paindlikult liigendatud selgroolülidest.
Konna koljus on vähem luid kui kala koljus. Kopsuhingamise tõttu ei ole konnal lõpuseid.
Jäsemete luustik vastab nende jagunemisele kolmeks osaks ja on jäsemete vööde luude kaudu ühendatud selgrooga. Esijäsemete vöö — rinnaku, kaks varese luud, kaks rangluud Ja kaks tera- on kaare välimusega ja paikneb lihaste paksuses. Tagajäsemete vöö moodustatud sulatatud vaagna luud ja on tihedalt selgroo külge kinnitatud. See toimib tagajäsemete toena.
Konna sisemine struktuur
Konna lihased
Konna lihaskonna struktuur on palju keerulisem kui kalal. Konn ju mitte ainult ei uju, vaid liigub ka maal. Lihaste või lihasrühmade kontraktsioonide kaudu saab konn teha keerulisi liigutusi. Tema jäsemelihased on eriti hästi arenenud.
Konna seedesüsteem
Kahepaiksete seedesüsteem on peaaegu sama ehitusega kui kaladel. Erinevalt kalast ei avane tema tagasool otse väljapoole, vaid spetsiaalsesse pikendusse, mida nimetatakse kloaak. Kloaaki avanevad ka suguelundite kusejuhad ja erituskanalid.

Joonis: Konna siseehitus. Konna seedesüsteem
Konna hingamiselundkond
Konn hingab atmosfääriõhku. Hingamiseks kasutatakse kopse ja nahka. Kopsud näevad välja nagu kotid. Nende seinad sisaldavad suurt hulka veresooni, milles toimub gaasivahetus. Konna kõri tõmmatakse mitu korda sekundis alla, luues suuõõnes haruldase ruumi. Seejärel tungib õhk läbi ninasõõrmete suuõõnde ja sealt edasi kopsudesse. See lükatakse tagasi keha seinte lihaste toimel. Konna kopsud on halvasti arenenud ja nahahingamine on tema jaoks sama oluline kui kopsuhingamine. Gaasivahetus on võimalik ainult siis, kui nahk on niiske. Kui konn panna kuiva anumasse, kuivab ta nahk peagi ära ja loom võib surra. Vette kastetuna lülitub konn täielikult nahahingamisele.

Joonis: Konna siseehitus. Konna vereringe- ja hingamissüsteemid
Konna vereringesüsteem
Konna süda asub keha esiosas, rinnaku all. See koosneb kolmest kambrist: vatsakese Ja kaks koda. Nii kodade kui ka seejärel vatsakese tõmbuvad kokku vaheldumisi.
Konna südames sisaldab parempoolne aatrium ainult venoosne veri, vasakule - ainult arteriaalne, ja vatsakeses on veri teatud määral segunenud.
Vatsakesest pärinevate veresoonte eriline paigutus viib selleni, et puhta arteriaalse verega varustatakse ainult konna aju, samas kui kogu keha saab segaverd.
Konna puhul voolab veri südame vatsakesest arterite kaudu kõikidesse organitesse ja kudedesse ning neist veenide kaudu paremasse aatriumisse - see süsteemne vereringe. Lisaks voolab veri vatsakesest kopsudesse ja nahka ning kopsudest tagasi südame vasakusse aatriumi – see kopsuvereringe. Kõigil selgroogsetel, välja arvatud kaladel, on kaks vereringeringi: väikesed - südamest hingamisorganitesse ja tagasi südamesse; suur - südamest arterite kaudu kõikidesse organitesse ja neist tagasi südamesse.
Ainevahetus kahepaiksetel konnade näitel
Ainevahetus kahepaiksetel on aeglane. Konna kehatemperatuur sõltub ümbritsevast temperatuurist: sooja ilmaga ta tõuseb ja külma ilmaga langeb. Kui õhk muutub väga kuumaks, langeb konna kehatemperatuur nahast niiskuse aurustumise tõttu. Nagu kalad, on ka konnad ja teised kahepaiksed külmaverelised loomad. Seetõttu muutuvad konnad külmemaks muutudes passiivseks, püüavad jõuda kuhugi soojemasse kohta ja talvel lähevad talveunne.
Kahepaiksete kesknärvisüsteem ja meeleelundid konna näitel
Kahepaiksete kesknärvisüsteem ja meeleelundid koosnevad samadest osadest, mis kaladel. Eesaju on arenenum kui kaladel ja selles saab eristada kahte turset - ajupoolkerad. Kahepaiksete kehad on maapinna lähedal ja nad ei pea tasakaalu hoidma. Sellega seoses on liigutuste koordineerimist kontrolliv väikeaju neil vähem arenenud kui kaladel.

Joonis: Konna siseehitus. Konna närvisüsteem
Meeleelundite ehitus vastab maapealsele keskkonnale. Näiteks silmalauge pilgutades eemaldab konn silma külge jäänud tolmuosakesed ja niisutab silma pinda.
Nagu kalal, on ka konnal sisekõrv. Õhus levivad helilained aga palju halvemini kui vees. Seetõttu on parema kuulmise huvides arenenud ka konn keskkõrv. See algab heli vastuvõtvast kuulmekilest, mis on õhuke ümmargune membraan silma taga. Sellest kanduvad helivõnked läbi kuulmisluu sisekõrva.
Loeng lisatud 02.05.2014 kell 10:01:44
Konna seedesüsteem koosneb suust, neelust, söögitorust, maost ja soolestikust. Konn püüab saaki kinni kleepuva keele abil, mis kinnitub esiotsast suu külge. Konn neelab kinnipüütud toidu tervelt alla. Konnadel on hästi arenenud magu, soolestikus on silmapaistvad kaksteistsõrmiksool, peen- ja jämesool. Maksa kanalid avanevad kaksteistsõrmiksoole koos pankrease kanaliga. Jämesool lõpeb pärasoolega, mis avaneb spetsiaalseks pikenduseks. nimetatakse kloaagiks.
Slaid 17 esitlusest "Konnade liigid". Arhiivi suurus koos esitlusega on 2385 KB.
Laadige esitlus alla
Bioloogia 8. klass
"Luumurrud" - orgaanilised ained - 60%. Uurige, milliseid omadusi anorgaanilised ained luudele annavad. Seejärel peseme luu. Alles jäid ainult anorgaanilised (mineraalsed) ained. Miks murduvad vanemad inimesed kukkudes luud tõenäolisemalt? Õppinud kirjandust uurides: 1. katse! Selle katlakivist eemaldatud luu saab sõlme siduda. Soovitused! Teeme järeldused! Kirjandusest saime teada, et luud sisaldavad: Kõik luu moodustavad orgaanilised ained põletatakse.
“Loomade ja taimede maailm” - vanad teod. Landrail. Riketid. Öökull. Vanker. Vutt. Taimed. Ööbikud. Tumbleweed. Harilik mutikriket. Rukis. Spurge. Putukad. Mürakas. Rohutirtsud. Liblikas. Lohe.
Mitu tiraaži on konnal?
Linnud. Loomad A.P. Tšehhovi "Stepp". Medvedka. Loomad. Little Bustard. Partridge. Gopher. Kanep.
"Inimese keha areng" - sperma sisemise struktuuri skeem. 8-rakuline embrüo. Sünni saladused. Blastula. Mis on sugurakkude põhiomadus? Inimese sünd. Sperma. Embrüo. 8 nädalat. Samara RCDO. Sperma kromosoomide komplekt. Muna. Embrüo. 5 nädalat. Embrüonaalsed rakud. 6 nädalat. Teine purustamine. Millised elundid arenevad esimesena? Kümnerakuline inimese embrüo.
"Punase raamatu linnud" - haud on levinud kõigil mandritel. Linnud. Samoilovski rajooni punast raamatut veel ei eksisteeri. kalakotkas. Mütsikute arvukus on kõvasti vähenenud. Väike tüübik on kana suurune. Öökull. kärbseseen. Linnud punasest raamatust. Little Bustard. Öökull on suuruse järgi kergesti äratuntav. Täiskasvanud lindudel on valge sulestik. Luik. Suur röövlind.
“Bioloogia “Inimese skelett”” - Skelett (skeletid - kuivatatud) - kõvade kudede kogum. Periost on luu ülemine kiht. Esiosa luu. Inimese luustik. Luu (os, ossis) on elund, selgroogse luustiku põhielement. Valige ainete loendist (1-10) õiged vastused küsimustele (A-M). Inimese luustikul on mitmeid erinevusi imetajate skeletist. Punane luuüdi on pehme kude. Skeleti lõigud. Rinnakorv. Rind on laienenud allapoole ja külgedele.
"Hingamisteede haigused ja vigastused" - bronhiidi sümptomid. Kopsupõletiku ennetamine. Kopsupõletik. Kopsupõletiku ravi. Bronhiit. Suitsetamine. Hingamisteede haigused. Kopsuvähi sümptomid ja põhjused. Suitsetamise mõju kopsudele. Nohu sümptomid. Bronhiidi ennetamine. Kopsuvähi ravi. Mittesuitsetaja kopsud. Bronhiidi ravi. Nohu ravi. Nohu. Nohu ennetamine. Hingamisteede haigused ja vigastused. Kopsud ja nende struktuur. Kopsupõletiku sümptomid.
Teemas “8. klassi bioloogia” on kokku 98 ettekannet
5klass.net > Bioloogia 8. klass > Konnade tüübid > Slaid 17
Veri täidab paljusid funktsioone ainult siis, kui see liigub läbi veresoonte. Ainevahetus vere ja teiste kehakudede vahel toimub kapillaaride võrgus. Seda eristab oma suur pikkus ja hargnevus, mis tagab suurepärase vastupidavuse verevoolule.
Veresoonte vastupanu ületamiseks vajalikku survet tekitab peamiselt süda Kalade südame ehitus on lihtsam kui kõrgematel selgroogsetel. Südame jõudlus survepumbana on kaladel oluliselt väiksem kui maismaaloomadel. Sellegipoolest tuleb ta oma ülesannetega toime. Veekeskkond loob soodsad tingimused südame toimimiseks. Kui maismaaloomadel kulub märkimisväärne osa südametööst gravitatsioonijõudude ja vere vertikaalliikumise ületamiseks, siis kalade puhul neutraliseerib tihe veekeskkond gravitatsioonimõjusid oluliselt. Horisontaalselt piklik keha, väike veremaht ja ainult ühe tsirkulatsiooniringi olemasolu hõlbustavad lisaks kalade südame talitlust.
Kala südame struktuur
Kala süda on väike, moodustades umbes 0,1% kehamassist. Sellest reeglist on muidugi erandeid. Näiteks lendkaladel ulatub südame mass 2,5%-ni kehakaalust.
Kõigil kaladel on kahekambriline süda. Selle elundi struktuuris on aga liigilisi erinevusi. Üldiselt võib kalade klassis ette kujutada kahte skeemi südame ehituse kohta. Nii esimesel kui ka teisel juhul eristatakse 4 õõnsust: venoosne siinus, aatrium, vatsake ja soojaverelistel loomadel ähmaselt aordikaare meenutav moodustis - teleostidel arterioossibul ja elastoharudel arteriooskoonus (joon. 7.1). Põhiline erinevus nende skeemide vahel seisneb vatsakeste ja arteriaalsete moodustiste morfofunktsionaalsetes omadustes.
Riis. 7.1. Kala südame ehituse skeem
Kala südame vatsakeses leiti erinevusi müokardi struktuuris. On üldtunnustatud seisukoht, et kalade müokard on spetsiifiline ja seda esindab homogeenne südamekude, millesse on ühtlaselt tunginud trabeekulid ja kapillaarid. Kalade lihaskiudude läbimõõt on väiksem kui soojaverelistel ja on 6-7 mikronit, mis on poole väiksem kui näiteks koera südamelihasel. Sellist müokardit nimetatakse käsnjaks. Teated kalade müokardi vaskularisatsiooni kohta on üsna segased. Müokard varustatakse venoosse verega trabekulaarsetest õõnsustest, mis omakorda täidetakse Teebea veresoonte kaudu vatsakesest verega. Klassikalises mõttes ei ole kaladel koronaarset vereringet. Vähemalt kardioloogid järgivad seda seisukohta. Siiski leidub ihtüoloogiat käsitlevas kirjanduses sageli mõistet "kalade koronaarringlus". Viimastel aastatel on teadlased avastanud palju variatsioone müokardi vaskularisatsioonis. Näiteks S. Agnisola et. al (1994) teatasid kahekihilise müokardi olemasolust forellil ja elektrikiirtel. Endokardi küljel asetseb käsnjas kiht ja selle kohal kompaktse, korrastatud paigutusega müokardi kiudude kiht.
Uuringud on näidanud, et müokardi käsnjas kiht varustatakse venoosse verega trabekulaarsetest lünkadest ja kompaktne kiht saab teise paari hargnevate pustulite hüpobronhiaalsete arterite kaudu arteriaalset verd. Elasmobranchide korral erineb koronaarvereringe selle poolest, et hüpobronhiaalsetest arteritest arteriaalne veri jõuab hästi arenenud kapillaarsüsteemi kaudu käsnjas kihti ja siseneb Tibesiuse veresoonte kaudu vatsakeste õõnsusse. Teine oluline erinevus teleostide ja elasmoharude vahel on perikardi morfoloogia.
Kala südame elektrilised omadused
Riis. 7.2. Kalade elektrokardiogramm
Forellil ja angerjal on elektrokardiogrammil selgelt näha lained P, Q, R, S ja T. Ainult S-laine näib hüpertrofeerunud ja Q-lainel on ootamatult positiivne suund, elasmoharudel lisaks viiele klassikalisele hambale. , näitab elektrokardiogramm Bd-laineid S ja hammaste T vahel, samuti Bg-hammast G- ja R-hammaste vahel. Angerja elektrokardiogrammil eelneb P lainele laine V. Lainete etioloogia on järgmine: P laine vastab kuulmekäigu ergastumisele ning venoosse siinuse ja aatriumi kokkutõmbumisele; QRS-kompleks iseloomustab atrioventrikulaarse sõlme ja vatsakeste süstoli ergastamist; T-laine tekib vastusena südame vatsakese rakumembraanide repolarisatsioonile.
Kala südame töö
Karpkala pulss (lööke minutis) temperatuuril 20 °C
Noorloomad kaaluga 0,02 g 80
Sõrmed kaaluvad 25 g 40
Kaheaastased kaaluga 500 g 30
In vitro katsetes (isoleeritud perfuseeritud süda) oli vikerforelli ja elektrikiire südame löögisagedus lööki minutis.
Kindlaks on tehtud kalade liigitundlikkus temperatuurimuutuste suhtes. Seega lesta puhul, kui vee temperatuur tõuseb g-lt 12 °C-ni, suureneb pulss 2 korda (24-lt 50 löögile minutis), ahvenal - ainult 30-36 lööki minutis.
Südame kontraktsioonide reguleerimine toimub kesknärvisüsteemi, aga ka intrakardiaalsete mehhanismide abil. Nagu soojaverelistel loomadel, täheldati ka kaladel in vivo katsetes tahhükardiat, kui südamesse voolava vere temperatuur tõusis. Südamesse voolava vere temperatuuri langus põhjustas bradükardia. Vagotoomia vähendas tahhükardia taset. Paljudel humoraalsetel teguritel on ka kronotroopne toime. Positiivne kronotroopne toime saavutati atropiini, adrenaliini ja eptatretiini manustamisega. Negatiivse kronotroopia põhjustasid atsetüülkoliin, efedriin ja kokaiin.
Huvitav on see, et sama humoraalne aine võib erinevatel ümbritseva õhu temperatuuridel avaldada kala südamele täpselt vastupidist mõju. Seega põhjustab epinefriin isoleeritud forelli südamel madalatel temperatuuridel (6°C) positiivset kronotroopset toimet ja perfusioonivedeliku kõrgendatud temperatuuride (15°C) taustal negatiivset kronotroopset toimet.
Kalade südame veretoodang on hinnanguliselt vml/kg minutis. Vere lineaarne kiirus kõhuaordis on cm/s. In vitro forellil tuvastati südame väljundi sõltuvus perfusioonivedeliku rõhust ja hapnikusisaldusest selles. Kuid samadel tingimustel ei muutunud elektrilise nõela minutimaht. Teadlased sisaldavad perfusaadis rohkem kui tosinat komponenti.
Naatriumkloriid 7,25
Kaaliumkloriid 0,23
Kaltsiumfluoriid 0,23
1. Lahus küllastatakse gaasiseguga, mis koosneb 99,5% hapnikust, 0,5% süsinikdioksiidist (süsinikdioksiid) või õhu (99 5%) ja süsinikdioksiidi (0,5%) seguga.
2. Perfusaadi pH reguleeritakse 7,9-ni temperatuuril 10 °C, kasutades naatriumvesinikkarbonaati.
Naatriumkloriid 16,36
Kaaliumkloriid 0,45
Magneesiumkloriid 0,61
Naatriumsulfaat 0,071
Naatriumvesinikkarbonaat 0,64
Kalade ringlus
Riis. 7.3. Luiste kalade vereringe diagramm
Unearterid ulatuvad eferentsetest haruarteritest peani. Järgmisena ühinevad haruarterid, moodustades ühtse suure veresoone - seljaaordi, mis ulatub kogu kehas selgroo alla ja tagab arteriaalse süsteemse vereringe. Peamised lahkuvad arterid on subklavia-, mesenteriaal-, niude-, saba- ja segmentaalarterid. Ringi venoosne osa algab lihaste ja siseorganite kapillaaridega, mis ühinevad, moodustades paaritud eesmised ja paaritud tagumised kardinaalsed veenid. Kardinaalveenid ühinevad kahe maksaveeniga, moodustades Cuvier' kanalid, mis tühjenevad siinusveeni.
Seega kala süda pumpab ja imeb ainult venoosset verd. Kuid
kõik elundid ja koed saavad arteriaalset verd, kuna enne elundite mikroveresoonkonna täitmist läbib veri lõpuseaparaadi, milles toimub gaasivahetus venoosse vere ja veekeskkonna vahel.
Vere liikumine ja vererõhk kaladel
Lisaks südamele aitavad veres veresoontes kaasa ka muud mehhanismid. Seega pakub dorsaalne aort, millel on suhteliselt jäikade (võrreldes kõhuaordiga) seintega sirge toru kuju, verevoolule vähe vastupanu. Segmentaalsetel, kaudaalsetel ja muudel arteritel on taskuklappide süsteem, mis sarnaneb suurte venoossete veresoonte omadega. See klapisüsteem takistab vere tagasivoolu. Venoosse verevoolu jaoks on suur tähtsus ka hiire veenidega külgnevatel kontraktsioonidel, mis suruvad verd südame suunas. Venoosne tagasivool ja südame väljund on optimeeritud salvestatud vere mobiliseerimisega. Eksperimentaalselt on tõestatud, et forellil põhjustab lihaskoormus põrna ja maksa mahu vähenemist. Lõpuks soodustab vere liikumist südame ühtlase täitumise mehhanism ja südame väljundi teravate süstool-diastoolsete kõikumiste puudumine. Südame täitumine on tagatud juba ventrikulaarse diastoli ajal, mil perikardiõõnde tekib mõningane vaakum ning veri täidab passiivselt venoosset siinust ja aatriumi. Süstoolset šokki summutab bulbus arteriosus, millel on elastne ja poorne sisepind.
Aqualover
Akvaariumid - akvaarium algajatele, akvaarium amatööridele, akvaarium professionaalidele
Peamenüü
Postituse navigeerimine
Kalade vereringesüsteem. Hematopoeetilised ja vereringeelundid
Enamus loetud
Külmaverelised (kehatemperatuur sõltub ümbritseva õhu temperatuurist) loomadel, kaladel, on suletud vereringesüsteem, mida esindavad süda ja veresooned. Erinevalt kõrgematest loomadest on kaladel üks tsirkulatsioon (erandiks on kopskalad ja labauimkalad).
Kala süda on kahekambriline: see koosneb aatriumist, vatsakesest, siinusvenosusest ja arterioossest koonusest, mis tõmbuvad vaheldumisi lihaste seintega kokku. Rütmiliselt kokku tõmbudes liigutab see verd nõiaringis.
Võrreldes maismaaloomadega on kala süda väga väike ja nõrk. Selle mass ei ületa tavaliselt 0,33–2,5%, keskmiselt 1% kehakaalust, imetajatel aga 4,6% ja lindudel 10–16%.
Kalade vererõhk on samuti nõrk.
Kaladel on ka madal pulss: 18–30 lööki minutis, kuid madalal temperatuuril võib see langeda 1–2-ni; Talvel jääks külmumise üle elanud kaladel lakkab sel perioodil südamepulss sootuks.
Lisaks on kaladel võrreldes kõrgemate loomadega väike kogus verd.
Kuid kõik see on seletatav kalade horisontaalse asendiga keskkonnas (pole vaja verd ülespoole suruda), aga ka kala eluga vees: keskkonnas, kus gravitatsioonijõud mõjutab palju. vähem kui õhus.
Veri voolab südamest arterite kaudu ja südamesse veenide kaudu.
Aatriumist surutakse see vatsakesesse, seejärel arteriosesse ja seejärel suurde kõhuaordi ning jõuab lõpustesse, kus toimub gaasivahetus: lõpuste veri rikastub hapnikuga ja vabaneb süsihappegaasist. Kalade punased verelibled – erütrotsüüdid – sisaldavad hemoglobiini, mis seob lõpustes hapnikku ning elundites ja kudedes süsihappegaasi.
Kalade veres oleva hemoglobiini võime hapnikku eraldada on liikide lõikes erinev. Kiiresti ujuvatel kaladel, kes elavad hapnikurikkas voolavas vees, on hemoglobiinirakud, millel on suurepärane hapniku sidumisvõime.
Hapnikurikkal arteriaalsel verel on helepunane värvus.
Pärast lõpuseid siseneb veri arterite kaudu pähe ja sealt edasi seljaaordi. Seljaaordi kaudu toimetab veri hapnikku kehatüve ja saba organitesse ja lihastesse. Sabaotsani ulatub seljaaort, millest mööda teed ulatuvad suured veresooned siseorganiteni.
Kalade veeniveri, hapnikuvaene ja süsihappegaasiga küllastunud, on tumeda kirsivärviga.
Olles andnud organitele hapnikku ja kogunud süsinikdioksiidi, voolab veri suurte veenide kaudu südamesse ja aatriumisse.
Kala kehal on ka vereloomes oma omadused:
Verd võivad moodustada paljud elundid: lõpuseaparaat, sooled (limaskest), süda (epiteelikiht ja veresoonte endoteel), neerud, põrn, veresoonte veri, lümfoidorgan (vereloome koe kogunemine – retikulaarne süntsüüt – kolju katuse all).
Kalade perifeerne veri võib sisaldada küpseid ja noori punaseid vereliblesid.
Erinevalt imetajate verest on punalibledel tuum.
Kala verel on sisemine osmootne rõhk.
Praeguseks on loodud 14 kalade veregrupisüsteemi.
Kellel on mitu vereringeringi?
Kahepaiksetel on kaks vereringeringi.
Imetajatel on kaks vereringet. Kahe ringi olemasolu tõttu vereringesüsteemis (väike ja suur) koosneb süda kahest osast: parempoolsest, mis pumpab verd väikesesse ringi, ja vasakpoolsest, mis väljutab verd suurde ringi. Vasaku vatsakese lihasmass on ligikaudu neli korda suurem kui parema lihasmass, mis on tingitud süsteemse ringi oluliselt suuremast vastupanuvõimest, kuid muud struktuurse korralduse tunnused on peaaegu identsed.
Rasedatel naistel on 3 ringi. Raseduse ajal täidab see süsteem topeltkoormust, kuna kehasse ilmub tegelikult "teine süda" - lisaks olemasolevale kahele vereringeringile moodustub vereringes uus lüli: nn emakas. platsenta verevool. Iga minut läbib selle ringi umbes 500 ml verd.
Raseduse lõpus suureneb veremaht kehas 6,5 liitrini. Selle põhjuseks on täiendava vereringe tekkimine, mis on loodud vastama loote kasvavatele vajadustele toitainete, hapniku ja ehitusmaterjalide järele.
Lülijalgsetel on avatud vereringesüsteem, mis tähendab, et vereringeringe pole.
Kaladel on üks vereringe ring.
Täiskasvanud kahepaiksetel on kaks vereringet.
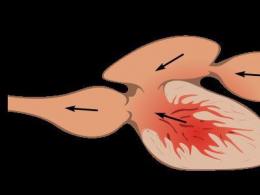
Selgroogsete vereringesüsteemid (kompleksid)
Kala südames on 4 järjestikku ühendatud õõnsust: sinus venosus, aatrium, vatsakese ja arteriooskoonus/kolb.
- Venoosne siinus (sinus venosus) on lihtne veeni pikendus, mis võtab verd.
- Haidel, ganoididel ja kopsukaladel sisaldab arteriooskoonus lihaskudet, mitut klappi ja on võimeline kokku tõmbuma.
- Luukaladel on arteriooskoonus vähenenud (ei ole lihaskudet ja klappe), seetõttu nimetatakse seda "arterikolbiks".
Kala südames olev veri on venoosne, sibulast/koonusest voolab see lõpustesse, seal muutub see arteriaalseks, voolab keha organitesse, muutub venoosseks, pöördub tagasi venoossesse siinusesse.
Kopsikala
Kopsukaladel ilmneb “kopsuvereringe”: viimasest (neljandast) lõpusearterist voolab veri läbi kopsuarteri (PA) hingamiskotti, kus see rikastub lisaks hapnikuga ja naaseb kopsuveeni (PV) kaudu tagasi süda, aatriumi vasakusse ossa. Kehast väljuv venoosne veri voolab, nagu peab, venoossesse siinusesse. Et piirata “kopsuringist” tuleva arteriaalse vere segunemist kehast väljuva venoosse verega, on aatriumis ja osaliselt vatsakeses mittetäielik vahesein.
Seega ilmub arteriaalne veri vatsakeses enne venoosset verd ja siseneb seetõttu eesmistesse haruarteritesse, kust viib otsetee pähe. Nutikas kalaaju saab kolm korda järjest gaasivahetusorganeid läbinud verd! Hapnikus suplemine, kelm.
Kahepaiksed
Kulleste vereringesüsteem on sarnane kondise kala omaga.
Täiskasvanud kahepaikse aatrium on vaheseinaga jagatud vasak- ja parempoolseks, mille tulemuseks on kokku 5 kambrit:
1) Kahepaiksete vasak aatrium saab arteriaalset verd kopsudest ja parem aatrium veeniverd elunditest ja arteriaalne veri nahast, seega konnade paremas aatriumis veri seguneb.
2) Nagu jooniselt näha, on arteriooskoonuse suu nihkunud parema aatriumi poole, nii et veri paremast aatriumist siseneb sinna esimesena ja vasakult - viimasena.
3) Arterioosi koonuse sees on spiraalklapp, mis jaotab kolm osa verd:
- esimene osa verd (paremast aatriumist, kõige venoossem) läheb naha kopsuarterisse (pulmokutaansesse arterisse) hapnikuga varustamiseks
- teine osa verd (segu paremast aatriumist ja arteriaalsest verest vasakust aatriumist) läheb süsteemse arteri kaudu kehaorganitesse
- kolmas osa verest (vasakust aatriumist, kõige arteriaalsem) läheb unearterisse (unearterisse) ajju.
4) Madalamatel kahepaiksetel (saba- ja jalgadeta) kahepaiksetel
- kodade vaheline vahesein on ebatäielik, seega toimub arteriaalse ja segavere segunemine tugevamalt;
- nahka varustatakse verega mitte naha kopsuarteritest (kus on võimalik kõige rohkem venoosset verd), vaid dorsaalsest aordist (kus veri on keskmine) - see ei ole eriti kasulik.
5) Kui konn istub vee all, voolab venoosne veri kopsudest vasakusse aatriumisse, mis teoreetiliselt peaks minema pähe. On olemas optimistlik versioon, et süda hakkab töötama erineval režiimil (muutub vatsakese ja arteriaalse koonuse pulsatsioonifaaside suhe), toimub vere täielik segunemine, mille tõttu ei satu kopsudest täielikult venoosne veri. pea, kuid segaveri, mis koosneb vasaku aatriumi venoossest verest ja parema aatriumi segaverest. On veel üks (pessimistlik) versioon, mille kohaselt saab veealuse konna aju kõige rohkem venoosset verd ja muutub tuhmiks.
Roomajad
Roomajate puhul väljuvad kopsuarter ("kopsu") ja kaks aordikaare vatsakesest, mis on osaliselt jagatud vaheseinaga. Vere jagunemine nende kolme veresoone vahel toimub samamoodi nagu kopsukaladel ja konnadel:
- Kõige rohkem arteriaalset verd (kopsudest) siseneb parempoolsesse aordikaare. Laste õppimise hõlbustamiseks algab aordi parempoolne kaar vatsakese vasakust osast ja seda nimetatakse "paremaks kaareks", kuna olles ümber südame paremale liikunud, kuulub see seljaaju arter (näete, kuidas see järgmisel ja järgnevatel joonistel välja näeb). Unearterid väljuvad paremast kaarest – kõige rohkem arteriaalset verd siseneb pähe;
- segaveri siseneb vasakusse aordikaare, mis läheb ümber südame vasakult ja ühendub parema aordikaarega - saadakse seljaajuarter, mis kannab verd organitesse;
- Kõige rohkem venoosset verd (kehaorganitest) siseneb kopsuarteritesse.
Krokodillid
Krokodillidel on neljakambriline süda, kuid nad segavad siiski verd läbi spetsiaalse Panizza avause vasaku ja parema aordikaare vahel.
Arvatakse aga, et tavaliselt segunemist ei toimu: kuna vasakus vatsakeses on kõrgem rõhk, ei voola sealt veri mitte ainult paremasse aordikaare (Parem aort), vaid ka läbi aordiava. Paanika - vasakusse aordikaare (vasak aort), seega saavad krokodilli elundid peaaegu täielikult arteriaalset verd.
Kui krokodill sukeldub, väheneb verevool läbi tema kopsude, rõhk paremas vatsakeses suureneb ja verevool läbi paanikaavade seiskub: veealuse krokodilli vasak aordikaar voolab paremast vatsakesest verd. Ma ei tea, mis mõte sellel on: kogu vereringesüsteemis olev veri on praegu venoosne, miks peaks seda kuhugi ümber jaotama? Igal juhul satub veri veealuse krokodilli pähe paremast aordikaarest – kui kopsud ei tööta, on see üleni venoosne. (Midagi ütleb mulle, et pessimistlik versioon kehtib ka veealuste konnade kohta.)
Linnud ja imetajad
Loomade ja lindude vereringesüsteemid kooliõpikutes on toodud tõele väga lähedal (nagu nägime, kõigil teistel selgroogsetel sellega nii palju ei veda). Ainus pisiasi, millest koolis rääkida ei tohi, on see, et imetajatel (B) säilib ainult vasak aordikaar ja lindudel (B) ainult parem (tähe A all on vereringesüsteem roomajatest, milles on arenenud mõlemad kaared) - Kanade ega inimeste vereringesüsteemis pole midagi muud huvitavat. Välja arvatud puuviljad...
Puuviljad
Arteriaalne veri, mille loote saab emalt, tuleb platsentast nabaveeni kaudu. Osa sellest verest siseneb maksa portaalsüsteemi, osa möödub maksast, mõlemad osad voolavad lõpuks alumisse õõnesveeni (sisemine õõnesveeni), kus segunevad looteorganitest voolava venoosse verega. Paremasse aatriumisse (RA) sisenedes lahjendatakse see veri veel kord ülemise õõnesveeni (ülemise õõnesveeni) veeniverega, mille tulemuseks on lootusetult segatud veri paremas aatriumis. Samal ajal satub osa veeniverd mittetöötavatest kopsudest loote vasakusse aatriumi – täpselt nagu krokodill istub vee all. Mida me teeme, kolleegid?
Appi tuleb vana hea mittetäielik vahesein, mille üle zooloogia kooliõpikute autorid nii kõvasti naeravad - inimese lootel, otse vasaku ja parema aatriumi vahelises vaheseinas, on ovaalne auk (Foramen ovale), mille kaudu siseneb paremast aatriumist segaveri vasakusse aatriumi. Lisaks on arterioosjuha (Dictus arteriosus), mille kaudu satub aordikaare paremast vatsakesest segatud veri. Seega voolab segaveri läbi loote aordi kõikidesse selle organitesse. Ja ka ajule! Ja sina ja mina kiusasime konni ja krokodille!! Ja nemad ise.
Testid
1. Kõhrekalade puudus:
a) ujumispõis;
b) spiraalklapp;
c) arteriooskoonus;
2. Imetajate vereringesüsteem sisaldab:
a) kaks aordikaare, mis seejärel ühinevad seljaaordiks;
b) ainult parempoolne aordikaar
c) ainult vasak aordikaar
d) ainult kõhuaort ja aordikaared puuduvad.
3. Lindude vereringesüsteem sisaldab:
A) kaks aordikaare, mis seejärel ühinevad seljaaordiks;
B) ainult parempoolne aordikaar;
B) ainult vasak aordikaar;
D) ainult kõhuaort ja aordikaared puuduvad.
4. Arteriaalne koonus on olemas
B) kõhrekalad;
D) kondine ganoidkala;
D) kondine kala.
5. Selgroogsete klassid, kus veri liigub otse hingamisorganitest keha kudedesse, ilma eelnevalt südant läbimata (valige kõik õiged valikud):
B) täiskasvanud kahepaiksed;
6. Kilpkonna süda oma struktuuris:
A) kolmekambriline mittetäieliku vaheseinaga vatsakeses;
D) neljakambriline, mille vatsakeste vahelises vaheseinas on auk.
7. Vereringe arv konnadel:
A) üks kullestel, kaks täiskasvanud konnadel;
B) üks täiskasvanud konnadel, kullestel puudub vereringe;
C) kaks kullestel, kolm täiskasvanud konnadel;
D) kaks kullestel ja täiskasvanud konnadel.
8. Selleks, et teie vasaku jala kudedest verre sattunud süsihappegaasi molekul saaks nina kaudu keskkonda sattuda, peab see läbima kõik teie keha järgmised struktuurid, välja arvatud:
B) kopsuveen;
B) kopsualveoolid;
D) kopsuarter.
9. Vereringel on kaks ringi (valige kõik õiged valikud):
A) kõhrelised kalad;
B) raisukala;
B) kopsukalad;
10. Neljakambrilisel südamel on:
11. Siin on skemaatiline joonis imetaja südamest. Hapnikuga veri siseneb südamesse järgmiste veresoonte kaudu:
12. Joonisel on kujutatud arterikaared:
Peatükk 7. KALADE VERERINGE OMADUSED
Veri täidab paljusid funktsioone ainult siis, kui see liigub läbi veresoonte. Ainevahetus vere ja teiste kehakudede vahel toimub kapillaaride võrgus. Seda eristab oma suur pikkus ja hargnevus, mis tagab suurepärase vastupidavuse verevoolule. Veresoonte takistuse ületamiseks vajaliku rõhu tekitab peamiselt süda,
Kalade südame ehitus on lihtsam kui kõrgematel selgroogsetel. Südame jõudlus survepumbana on kaladel oluliselt väiksem kui maismaaloomadel. Sellegipoolest tuleb ta oma ülesannetega toime. Veekeskkond loob soodsad tingimused südame toimimiseks. Kui maismaaloomadel kulub märkimisväärne osa südametööst gravitatsioonijõudude ja vere vertikaalliikumise ületamiseks, siis kalade puhul neutraliseerib tihe veekeskkond gravitatsioonimõjusid oluliselt. Horisontaalselt piklik keha, väike veremaht ja ainult ühe tsirkulatsiooniringi olemasolu hõlbustavad lisaks kalade südame talitlust.
§ kolmkümmend. SÜDAME STRUKTUUR
Kõigil kaladel on kahekambriline süda. Selle elundi struktuuris on aga liigilisi erinevusi. Üldiselt võib kalade klassis ette kujutada kahte skeemi südame ehituse kohta. Nii esimesel kui ka teisel juhul eristatakse 4 õõnsust: venoosne siinus, aatrium, vatsake ja soojaverelistel loomadel ähmaselt aordikaare meenutav moodustis - teleostidel arterioossibul ja elastoharudel arteriooskoonus (joon. 7.1).
Põhiline erinevus nende skeemide vahel seisneb vatsakeste ja arteriaalsete moodustiste morfofunktsionaalsetes omadustes.
Teleostides on arterikolb esindatud kiudkoega, millel on sisemise kihi käsnjas struktuur, kuid ilma ventiilideta.
Elasmoharudes sisaldab arteriooskoonus lisaks kiulisele koele ka tüüpilist südamelihaskoe ja seetõttu on tal kontraktiilsus. Koonusel on klappide süsteem, mis hõlbustab vere ühesuunalist liikumist läbi südame.
Kala südame vatsakeses leiti erinevusi müokardi struktuuris. On üldtunnustatud seisukoht, et kalade müokard on spetsiifiline ja seda esindab homogeenne südamekude, millesse on ühtlaselt tunginud trabeekulid ja kapillaarid. Kalade lihaskiudude läbimõõt on väiksem kui soojaverelistel ja on 6-7 mikronit, mis on poole väiksem kui näiteks koera südamelihasel. Sellist müokardit nimetatakse käsnjaks.
Teated kalade müokardi vaskularisatsiooni kohta on üsna segased. Müokard varustatakse venoosse verega trabekulaarsetest õõnsustest, mis omakorda täidetakse Teebea veresoonte kaudu vatsakesest verega. Klassikalises mõttes ei ole kaladel koronaarset vereringet. Vähemalt kardioloogid järgivad seda seisukohta. Siiski leidub ihtüoloogiat käsitlevas kirjanduses sageli mõistet "kalade koronaarringlus".
Viimastel aastatel on teadlased avastanud palju variatsioone müokardi vaskularisatsioonis. Näiteks S. Agnisola et. al (1994) teatasid kahekihilise müokardi olemasolust forellil ja elektrikiirtel. Endokardi küljel asetseb käsnjas kiht ja selle kohal kompaktse, korrastatud paigutusega müokardi kiudude kiht.
Uuringud on näidanud, et müokardi käsnjas kiht varustatakse venoosse verega trabekulaarsetest lünkadest ja kompaktne kiht saab teise paari hargnevate pustulite hüpobronhiaalsete arterite kaudu arteriaalset verd. Elasmobranchide korral erineb koronaarvereringe selle poolest, et hüpobronhiaalsetest arteritest arteriaalne veri jõuab hästi arenenud kapillaarsüsteemi kaudu käsnjas kihti ja siseneb Tibesiuse veresoonte kaudu vatsakeste õõnsusse.
Teine oluline erinevus teleostide ja elasmoharude vahel on perikardi morfoloogia.
Teleostidel meenutab perikardit maismaaloomade oma. Seda esindab õhuke kest.
Elasmoharudes moodustab perikardi kõhrekoe, seega on see nagu kõva, kuid elastne kapsel. Viimasel juhul tekib diastoli ajal perikardi ruumis teatav vaakum, mis hõlbustab verevarustust venoossesse siinusesse ja aatriumi ilma täiendava energiakuluta.
§31. SÜDAME ELEKTRILISED OMADUSED
Kalade südamelihase müotsüütide struktuur on sarnane kõrgemate selgroogsete omaga. Seetõttu on südame elektrilised omadused sarnased. Müotsüütide puhkepotentsiaal teleostidel ja elasmobranchidel on 70 mV, hakkkaladel 50 mV. Aktsioonipotentsiaali tipul registreeritakse potentsiaali märgi ja suuruse muutus miinus 50 mV-lt pluss 15 mV-ni. Müotsüütide membraani depolarisatsioon põhjustab naatrium-kaltsiumikanalite ergastamist. Esiteks tormavad müotsüütide rakku naatriumiioonid ja seejärel kaltsiumiioonid. Selle protsessiga kaasneb venitatud platoo moodustumine ja südamelihase absoluutne tulekindlus registreeritakse funktsionaalselt. See faas kaladel on palju pikem - umbes 0,15 s.
Järgnev kaaliumikanalite aktiveerimine ja kaaliumiioonide vabanemine rakust tagavad müotsüütide membraani kiire repolarisatsiooni. Membraani repolarisatsioon omakorda sulgeb kaaliumikanalid ja avab naatriumikanalid. Selle tulemusena taastub rakumembraani potentsiaal algsele tasemele miinus 50 mV.
Kala südame müotsüüdid, mis on võimelised genereerima potentsiaali, paiknevad teatud südamepiirkondades, mis on kollektiivselt ühendatud "südame juhtivaks süsteemiks". Nagu kõrgematel selgroogsetel, toimub ka kaladel südame süstool sünatriaalsõlmes.
Erinevalt teistest selgroogsetest täidavad kaladel südamestimulaatorite rolli kõik juhtivussüsteemi struktuurid, mis teleostidel hõlmab kõrvakanali keskpunkti, sõlme atrioventrikulaarses vaheseinas, millest Purkinje rakud ulatuvad vatsakese tüüpiliste kardiotsüüdideni. .
Ergastuse kiirus läbi südame juhtivuse on kaladel väiksem kui imetajatel ja see on südame eri osades erinev. Potentsiaalse leviku maksimaalne kiirus registreeriti vatsakese struktuurides.
Kalade elektrokardiogramm sarnaneb juhtmetes V3 ja V4 inimese elektrokardiogrammiga (joonis 7.2). Pliisid kalade puhul ei ole aga nii detailselt välja töötatud kui maismaaselgroogsete puhul.
Riis. 7.2. Kalade elektrokardiogramm
Forellil ja angerjal on elektrokardiogrammil selgelt näha lained P, Q, R, S ja T. Ainult S-laine näib hüpertrofeerunud ja Q-lainel on ootamatult positiivne suund, elasmoharudel lisaks viiele klassikalisele hambale. , näitab elektrokardiogramm Bd-laineid S ja hammaste T vahel, samuti Br-hammast G- ja R-hammaste vahel. Angerja elektrokardiogrammil eelneb P-lainele laine V. Lainete etioloogia on järgmine:
P-laine vastab kõrvakanali ergutusele ning venoosse siinuse ja aatriumi kokkutõmbumisele;
QRS-kompleks iseloomustab atrioventrikulaarse sõlme ja vatsakeste süstoli ergastamist;
T-laine tekib vastusena südame vatsakese rakumembraanide repolarisatsioonile.
Kala süda töötab rütmiliselt. Kalade pulss sõltub paljudest teguritest.
Südame löögisagedus (lööke minutis) karpkalal temperatuuril 20 °C
Noorloomad kaaluga 0,02 g 80
Sõrmed kaaluvad 25 g 40
Kaheaastased kaaluga 500 g 30
Paljudest teguritest mõjutab südame löögisagedust kõige rohkem keskkonna temperatuur. Kasutades telemeetria meetodit meriahven ja lesta puhul, selgus järgmine seos (tabel 7.1).
7.1. Südame löögisageduse sõltuvus vee temperatuurist
Kindlaks on tehtud kalade liigitundlikkus temperatuurimuutuste suhtes. Seega lesta puhul, kui vee temperatuur tõuseb g-lt 12 °C-ni, suureneb pulss 2 korda (24-lt 50 löögile minutis), ahvenal - ainult 30-36 lööki minutis.
Südame kontraktsioonide reguleerimine toimub kesknärvisüsteemi, aga ka intrakardiaalsete mehhanismide abil. Nagu soojaverelistel loomadel, täheldati ka kaladel in vivo katsetes tahhükardiat, kui südamesse voolava vere temperatuur tõusis. Südamesse voolava vere temperatuuri langus põhjustas bradükardia. Vagotoomia vähendas tahhükardia taset.
Paljudel humoraalsetel teguritel on ka kronotroopne toime. Positiivne kronotroopne toime saavutati atropiini, adrenaliini ja eptatretiini manustamisega. Negatiivse kronotroopia põhjustasid atsetüülkoliin, efedriin ja kokaiin.
Huvitav on see, et sama humoraalne aine võib erinevatel ümbritseva õhu temperatuuridel avaldada kala südamele täpselt vastupidist mõju. Seega põhjustab epinefriin isoleeritud forelli südamel madalatel temperatuuridel (6°C) positiivset kronotroopset toimet ja perfusioonivedeliku kõrgendatud temperatuuride (15°C) taustal negatiivset kronotroopset toimet.
Kalade südame veretoodang on hinnanguliselt vml/kg minutis. Vere lineaarne kiirus kõhuaordis on cm/s. In vitro forellil tuvastati südame väljundi sõltuvus perfusioonivedeliku rõhust ja hapnikusisaldusest selles. Kuid samadel tingimustel ei muutunud elektrilise nõela minutimaht.
Teadlased sisaldavad perfusaadis rohkem kui tosinat komponenti.
Forelli südame perfusaadi koostis (g/l)
Naatriumkloriid 7,25
Kaaliumkloriid 0,23
Kaltsiumfluoriid 0,23
Magneesiumsulfaat (kristalne) 0,23
Naatriumfosfaat monoasendatud (kristalne) 0,016
Dinaatriumfosfaat (kristalne) 0,41
Polüvinüülpürrooliidol (PVP) kolloidne 10.0
I. Lahus küllastatakse gaasiseguga, mis koosneb 99,5% hapnikust, 0,5% süsinikdioksiidist (süsinikdioksiid) või õhu segust (99 5%) ja süsinikdioksiidist (0,5%).
2. Perfusaadi pH reguleeritakse 7,9-ni temperatuuril 10 °C, kasutades naatriumvesinikkarbonaati.
Elektrilise kiirsüdame perfusaadi koostis (g/l)
Naatriumkloriid 16,36
Kaaliumkloriid 0,45
Magneesiumkloriid 0,61
Naatriumsulfaat 0,071
Naatriumfosfaat monoasendatud (kristalne) 0,14
Naatriumvesinikkarbonaat 0,64
1. Perfusaat küllastatakse sama gaasiseguga. 2.pH 7,6.
Sellistes lahustes säilitab isoleeritud kalasüda oma füsioloogilised omadused ja funktsioonid väga pikka aega. Südamega lihtsate manipulatsioonide tegemisel on lubatud kasutada isotoonilist naatriumkloriidi lahust. Siiski ei tohiks loota südamelihase pikaajalisele tööle.
Kaladel, nagu teate, on üks vereringe ring. Ja veel, veri ringleb läbi selle kauem. Kalal kulub täielikuks vereringeks umbes 2 minutit (inimesel läbib veri kahte vereringeringi). Vatsakesest bulbus arteriosuse ehk conus arteriosuse kaudu siseneb veri nn kõhuaordi, mis ulatub südamest kraniaalses suunas kuni lõpusteni (joon. 7.3).
Kõhuaort jaguneb vasak- ja parempoolseks (harukaarte arvu järgi) aferentseks haruarteriks. Nendest ulatub igale lõpuse filamendile kroonlehearter ja sellest iga kroonleheni väljub kaks arteriooli, mis moodustavad kõige peenemate veresoonte kapillaarvõrgu, mille seina moodustab ühekihiline epiteel, millel on suured rakkudevahelised ruumid. Kapillaarid ühinevad üheks eferentseks arteriooliks (vastavalt kroonlehtede arvule). Eferentsed arterioolid moodustavad efferentse kroonlehe arteri. Kroonlehearterid moodustavad vasaku ja parema eferentse haruarteri, mille kaudu voolab arteriaalne veri.
Riis. 7.3. Luude kalade vereringe diagramm:
1- kõhuaort; 2 - unearterid; 3 - lõpuste arterid; 4- subklavia arter ja veen; b- dorsaalne aort; 7 - tagumine kardinaalveen; 8- neerusooned; 9 - sabaveen; 10 - neerude pöörduv veen; 11 - soolestiku anumad, 12 - portaalveen; 13 - maksa veresooned; 14- maksa veenid; 15- venoosne 16- Cuvier' kanal; 17- eesmine kardinaalveen
Unearterid ulatuvad eferentsetest haruarteritest peani. Järgmisena ühinevad haruarterid, moodustades ühtse suure veresoone - seljaaordi, mis ulatub kogu kehas selgroo alla ja tagab arteriaalse süsteemse vereringe. Peamised lahkuvad arterid on subklavia-, mesenteriaal-, niude-, saba- ja segmentaalarterid.
Ringi venoosne osa algab lihaste ja siseorganite kapillaaridega, mis ühinevad, moodustades paaritud eesmised ja paaritud tagumised kardinaalsed veenid. Kardinaalveenid ühinevad kahe maksaveeniga, moodustades Cuvier' kanalid, mis tühjenevad siinusveeni.
Seega kala süda pumpab ja imeb ainult venoosset verd. Arteriaalset verd saavad aga kõik elundid ja koed, kuna enne elundite mikroveresoonkonna täitmist läbib veri lõpuseaparaadi, milles toimub gaasivahetus venoosse vere ja veekeskkonna vahel.
§34. VERE LIIKUMINE JA VERERÕHK
Veri liigub veresoontes selle rõhu erinevuse tõttu vereringe alguses ja lõpus. Kui vererõhku mõõdeti ilma anesteesiata ventraalses asendis (põhjustades bradükardiat), oli lõhe kõhuaordis 82/50 mm Hg. Art., ja seljaosas 44/37 mm Hg. Art. Mitme liigi tuimestatud kalade uuring näitas, et tuimestus alandas oluliselt süstoolset vererõhku – domm Hg. Art. Pulsirõhk varieerus kalaliikide lõikes 10–30 mm Hg. Art. Hüpoksia tõi kaasa pulsirõhu tõusu 40 mmHg-ni. Art.
Vereringe lõppedes ei ületanud vererõhk veresoonte seintel (Cuvier kanalites) 10 mm Hg. Art.
Suurima vastupanu verevoolule pakub lõpustesüsteem oma pikkade ja väga hargnenud kapillaaridega. Karpkalal ja forellil on süstoolse rõhu erinevus kõhu- ja seljaaordis ehk lõpuseaparaadi sisenemisel ja väljumisel %. Hüpoksia ajal pakuvad lõpused veelgi suuremat vastupanu verevoolule.
Lisaks südamele aitavad veres veresoontes kaasa ka muud mehhanismid. Seega pakub dorsaalne aort, millel on suhteliselt jäikade (võrreldes kõhuaordiga) seintega sirge toru kuju, verevoolule vähe vastupanu. Segmentaalsetel, kaudaalsetel ja muudel arteritel on taskuklappide süsteem, mis sarnaneb suurte venoossete veresoonte omadega. See klapisüsteem takistab vere tagasivoolu. Venoosse verevoolu jaoks on suur tähtsus ka hiire veenidega külgnevatel kontraktsioonidel, mis suruvad verd südame suunas.
Venoosne tagasivool ja südame väljund on optimeeritud salvestatud vere mobiliseerimisega. Eksperimentaalselt on tõestatud, et forellil põhjustab lihaskoormus põrna ja maksa mahu vähenemist.
Lõpuks soodustab vere liikumist südame ühtlase täitumise mehhanism ja südame väljundi teravate süstool-diastoolsete kõikumiste puudumine. Südame täitumine on tagatud juba ventrikulaarse diastoli ajal, mil perikardiõõnde tekib mõningane vaakum ning veri täidab passiivselt venoosset siinust ja aatriumi. Süstoolset šokki summutab bulbus arteriosus, millel on elastne ja poorne sisepind.
Hapniku kontsentratsioon veehoidlas on kalade elupaiga kõige ebastabiilsem näitaja, mis muutub päeva jooksul mitu korda. Sellest hoolimata on hapniku ja süsihappegaasi osarõhk kalade veres üsna stabiilne ja kuulub homöostaasi jäikade konstantide hulka.
Hingamiskeskkonnana on vesi halvem kui õhk (tabel 8.1).
8.1. Vee ja õhu kui hingamiskeskkonna võrdlus (temperatuuril 20 °C)
Arvestades nii ebasoodsaid gaasivahetuse algtingimusi, on evolutsioon võtnud teed, mis loovad veeloomadele täiendavaid gaasivahetusmehhanisme, mis võimaldavad neil taluda ohtlikke hapniku kontsentratsiooni kõikumisi oma keskkonnas. Lisaks kalade lõpustele osalevad gaasivahetuses nahk, seedetrakt, ujupõis ja spetsiaalsed elundid.
§35. LÕPED – VEESESKKONNAS EFEKTIIVNE GAASIVAHETUSE ORGAN
Peamine koorem kalade hapnikuga varustamise ja sealt süsihappegaasi eemaldamise eest langeb lõpustele. Nad teevad teetanilist tööd. Kui võrrelda lõpuse- ja kopsuhingamist, siis jõuate järeldusele, et kala peab läbi lõpuste pumpama hingamiskeskkonda, mis on mahult 30 korda suurem ja massilt (!) korda suurem.
Lähem uurimine näitab, et lõpused on hästi kohanenud gaasivahetuseks veekeskkonnas. Hapnik liigub lõpuste kapillaarikihti mööda osarõhugradienti, mis kaladel on mm Hg. Art. Sellest tuleneb ka hapniku üleminek verest kudedes rakkudevahelisele vedelikule.
Siin on hapniku osarõhu gradient hinnanguliselt 1–15 mmHg. Art., Süsinikdioksiidi kontsentratsiooni gradient - 3-15 mmHg.
Gaasivahetus teistes elundites, näiteks läbi naha, toimub samade füüsikaliste seaduste järgi, kuid difusiooni intensiivsus neis on palju väiksem. Lõpuse pind on mitu korda suurem kui kala kehapind. Lisaks on lõpustel, gaasivahetuseks väga spetsialiseerunud elunditel, suured eelised isegi teiste elunditega samas piirkonnas.
Lõpuseaparaadi kõige täiuslikum struktuur on iseloomulik kondistele kaladele. Lõpuseaparaadi aluseks on 4 paari lõpusekaari. Lõpusevõlvidel on hästi vaskulariseeritud lõpuse niidid, mis moodustavad hingamispinna (joon. 8.1).
Lõpusekaare suuõõne poole jääval küljel on väiksemad struktuurid – lõpusekaapad, mis vastutavad suures osas suuõõnest lõpuseniitidele voolava vee mehaanilise puhastamise eest.
Mikroskoopilised lõpuseniidid paiknevad põiki lõpuse filamentide suhtes, mis on lõpuste kui hingamiselundite struktuurielemendid (vt joon. 8.1; 8.2). Kroonlehti katvas epiteelis on kolme tüüpi rakke: hingamis-, limaskesta- ja toetavad rakud. Sekundaarsete lamellide pindala ja sellest tulenevalt ka hingamisteede epiteel sõltub kalade bioloogilistest omadustest - elustiilist, põhiainevahetuse kiirusest, hapnikuvajadusest. Niisiis on 100 g massiga tuunikala lõpuse pindala cm 2 / g, mulletil - 10 cm 2 / g, forellil - 2 cm 2 / g, särjel - 1 cm 2 / g.
Lõpusegaasivahetus saab olla efektiivne ainult siis, kui vesi voolab läbi lõpuseaparaadi. Vesi niisutab pidevalt lõpuse filamente ja seda hõlbustab suuline aparaat. Vesi tormab suust lõpustesse. Enamikul kalaliikidel on see mehhanism.
Riis. 8.1. Luukalade lõpuste ehitus:
1- lõpuse niidid; 2- lõpuse niidid; 3-haruline arter; 4 - haruline veen; 5-kroonlehe arter; 6 - kroonlehe veen; 7 lõpusega rehad; 8-lõheline kaar
Küll aga on teada, et suured ja aktiivsed liigid, näiteks tuunikala, ei sulge oma suud ning neil puuduvad lõpusekatete hingamisliigutused. Seda tüüpi lõpuste ventilatsiooni nimetatakse "oinaks"; see on võimalik ainult vees suurel liikumiskiirusel.
Vee läbimist lõpuste kaudu ja vere liikumist lõpuseaparaadi veresoontes iseloomustab vastuvoolumehhanism, mis tagab gaasivahetuse väga kõrge efektiivsuse. Pärast lõpuste läbimist kaotab vesi kuni 90% selles lahustunud hapnikust (tabel 8.2).
8.2. Veest hapniku eraldamise efektiivsus erinevate kalaharkidega, %
Lõpuseniidid ja kroonlehed paiknevad väga lähestikku, kuid neid läbiva vee väikese kiiruse tõttu ei tekita nad veevoolule erilist takistust. Arvutuste kohaselt on kalade energiakulud hoolimata suurest töömahust, mis on seotud vee liigutamisega läbi lõpuseaparaadi (vähemalt 1 m 3 vett 1 kg eluskaalu kohta ööpäevas).
Vee sissepritse tagavad kaks pumpa - suu ja lõpus. Erinevatel kalaliikidel võib üks neist domineerida. Näiteks kiiresti liikuvatel mulletel ja stauriididel töötab peamiselt suupump, aeglaselt liikuvatel põhjakaladel (lest või säga) aga nakkepump.
Hingamisliigutuste sagedus kaladel sõltub paljudest teguritest, kuid kõige rohkem mõjutavad seda füsioloogilist näitajat kaks - vee temperatuur ja hapnikusisaldus selles. Hingamissageduse sõltuvus temperatuurist on näidatud joonisel fig. 8.2.
Seega tuleks nakkehingamist pidada väga tõhusaks gaasivahetuse mehhanismiks veekeskkonnas nii hapniku eraldamise tõhususe kui ka selle protsessi energiatarbimise seisukohalt. Juhul, kui lõpusemehhanism ei suuda piisava gaasivahetuse ülesandega toime tulla, aktiveeritakse muud (abi)mehhanismid.
Nahahingamine on kõigil loomadel erineval määral arenenud, kuid mõne kalaliigi puhul võib see olla peamine gaasivahetuse mehhanism.
Nahahingamine on hädavajalik liikidele, kes elavad vähehapniku tingimustes istuvat eluviisi või lahkuvad reservuaarist lühikeseks ajaks (angerjas, mudakala, säga). Täiskasvanud angerjal muutub naha hingamine peamiseks ja ulatub 60% gaasivahetuse kogumahust.
8.3. Nahahingamise osakaal erinevatel kalaliikidel
Kalade ontogeneetilise arengu uuring näitab, et nahahingamine on lõpusehingamise suhtes esmane. Kalade embrüod ja vastsed vahetavad gaase keskkonnaga läbi sisekudede. Nahahingamise intensiivsus suureneb vee temperatuuri tõustes, kuna temperatuuri tõus suurendab ainevahetust ja vähendab hapniku lahustuvust vees.
Üldiselt määrab naha gaasivahetuse intensiivsuse naha morfoloogia. Angerjatel on nahal teiste liikidega võrreldes hüpertrofeerunud vaskularisatsioon ja innervatsioon.
Teistel liikidel, näiteks haidel, on nahahingamise osatähtsus tähtsusetu, kuid ka nende nahk on kareda struktuuriga koos halvasti arenenud verevarustussüsteemiga.
Nahaveresoonte pindala on erinevatel luukalaliikidel vahemikus 0,5–1,5 cm:/g eluskaalu kohta. Nahakapillaaride ja nakkekapillaaride pindala suhe on väga erinev - 3:1 säärtel kuni 10:1 karpkalal.
Epidermise paksus, mis ulatub 263 µm lesta kuni 263 µm angerjas ja 338 µm lehas, määratakse limaskestarakkude arvu ja suuruse järgi. Küll aga leidub kalu väga intensiivse gaasivahetusega naha tavalise makro- ja mikrostruktuuri taustal.
Kokkuvõtteks tuleb rõhutada, et loomade nahahingamise mehhanismi ei ole ilmselgelt piisavalt uuritud. Olulist rolli selles protsessis mängib naha lima, mis sisaldab nii hemoglobiini kui ka ensüümi karboanhüdraasi.
Ekstreemsetes tingimustes (hüpoksia) kasutavad soolehingamist paljud kalaliigid. Siiski on kalu, mille seedetraktis on efektiivse gaasivahetuse eesmärgil toimunud morfoloogilised muutused. Sel juhul reeglina soolestiku pikkus suureneb. Sellistes kalades (säga, kalja) neelatakse õhk alla ja suunatakse soolestiku peristaltiliste liigutustega spetsiaalsesse sektsiooni. Seedetrakti selles osas on sooleseinad kohandatud gaasivahetuseks esiteks hüpertrofeerunud kapillaaride vaskularisatsiooni tõttu ja teiseks hingamisteede kolonnepiteeli olemasolu tõttu. Allaneelatud atmosfääriõhu mull soolestikus on teatud rõhu all, mis suurendab hapniku difusioonikoefitsienti verre. Selles kohas varustatakse soolestikku venoosse verega, mistõttu on hea hapniku ja süsihappegaasi osarõhu erinevus ning nende difusiooni ühesuunalisus. Soolehingamine on Ameerika sägadel laialt levinud. Nende hulgas on liike, mille kõht on kohandatud gaasivahetuseks.
Ujumispõis ei anna kaladele mitte ainult neutraalset ujuvust, vaid mängib rolli ka gaasivahetuses. See võib olla avatud (lõhe) või suletud (karpkala). Avatud põis on õhukanali kaudu ühendatud söögitoruga ning selle gaasikoostist saab kiiresti uuendada. Suletud põies toimuvad muutused gaasi koostises ainult vere kaudu.
Ujumispõie seinas on spetsiaalne kapillaarsüsteem, mida tavaliselt nimetatakse "gaasinäärmeks". Nääre kapillaarid moodustavad järsult kõverdatud vastuvoolu silmuseid. Gaasinäärme endoteel on võimeline eritama piimhapet ja muutma seeläbi lokaalselt vere pH-d. See omakorda põhjustab hemoglobiini hapniku otse vereplasmasse eraldumise. Selgub, et ujupõiest voolav veri on hapnikuga üleküllastunud. Kuid gaasinäärme verevoolu vastuvoolumehhanism põhjustab selle plasma hapniku difundeerumist põieõõnde. Seega loob mull hapnikuvaru, mida kala keha kasutab ebasoodsates tingimustes.
Teisi gaasivahetuse seadmeid esindavad labürint (gourami, lalius, kukk), epibranhiaalne organ (riisiangerjas), kopsud (kopsukala), suuaparaat (roomakas), neeluõõnsused (Ophiocephalus sp.). Gaasivahetuse põhimõte nendes elundites on sama, mis soolestikus või ujupõies. Gaasivahetuse morfoloogiline alus neis on modifitseeritud kapillaarringlussüsteem pluss limaskestade hõrenemine (joon. 8.3).
1- liugur ahven: 2- hunnik; 3- ussipea; 4-Niiluse Sharmuth
Morfoloogiliselt ja funktsionaalselt on pseudobranchia - lõpuseaparaadi erilised moodustised - seotud hingamiselunditega. Nende rolli ei mõisteta täielikult. See. et hapnikuga küllastunud veri voolab nendesse struktuuridesse lõpustest viitab sellele. et nad ei osale hapnikuvahetuses. Suure koguse karboanhüdraasi olemasolu pseudobranchia membraanidel viitab aga nende struktuuride osalemisele lõpuseaparaadi süsinikdioksiidi vahetuse reguleerimises.
Funktsionaalselt on pseudobranchiaga ühendatud nn veresoonte nääre, mis asub silmamuna tagumisel seinal ja ümbritseb nägemisnärvi. Vaskulaarsel näärmel on kapillaaride võrgustik, mis meenutab ujupõie gaasinäärme võrgustikku. On seisukoht, et vaskulaarne nääre tagab silma võrkkesta kõrge hapnikusisaldusega verevarustuse võimalikult vähese süsihappegaasi sissevõtmisega. On tõenäoline, et fotoretseptsioon on nõudlik nende lahuste pH suhtes, milles see esineb. Seetõttu võib pseudobranhiaal-veresoonkonna näärmesüsteemi pidada võrkkesta täiendavaks puhverfiltriks. Kui võtta arvesse, et selle süsteemi olemasolu ei ole seotud kalade taksonoomilise asendiga, vaid pigem elupaigaga (need elundid esinevad sagedamini mereliikidel, kes elavad suure läbipaistvusega vees ja mille jaoks nägemus on kõige olulisem suhtluskanal väliskeskkonnaga) , siis tundub see eeldus veenev.
Kalades gaaside transportimisel verega põhimõttelisi erinevusi ei ole. Nagu kopsuloomadel, realiseeritakse ka kaladel vere transpordifunktsioonid tänu hemoglobiini kõrgele afiinsusele hapniku suhtes, gaaside suhteliselt suurele lahustuvusele vereplasmas ning süsinikdioksiidi keemilisele muundamisele karbonaatideks ja vesinikkarbonaatideks.
Peamine hapniku transportija kalade veres on hemoglobiin. Huvitav on see, et kalade hemoglobiin jaguneb funktsionaalselt kahte tüüpi – happetundlikuks ja happetundlikuks.
Happetundlik hemoglobiin kaotab vere pH langedes hapniku sidumise võime.
Happe suhtes tundetu hemoglobiin ei reageeri pH väärtustele ning kalade jaoks on selle olemasolu elulise tähtsusega, kuna nende lihaste aktiivsusega kaasneb suur piimhappe eraldumine verre (konstantse glükolüüsi loomulik tulemus). hüpoksia).
Mõnede Arktika ja Antarktika kalaliikide veres pole hemoglobiini üldse. Kirjanduses on teateid sama nähtuse kohta karpkala puhul. Forelliga tehtud katsed näitasid, et ilma funktsionaalse hemoglobiinita (kogu hemoglobiin seoti kunstlikult CO abil) ei esine kaladel lämbumist veetemperatuuril alla 5 °C. See näitab, et kalade hapnikuvajadus on oluliselt väiksem kui maismaaloomadel (eriti madalal veetemperatuuril, kui gaaside lahustuvus vereplasmas suureneb).
Teatud tingimustel tuleb gaaside transpordiga toime ainult plasma. Kuid tavatingimustes on valdavas enamuses kaladest gaasivahetus ilma hemoglobiinita praktiliselt välistatud. Hapniku difusioon veest verre toimub piki kontsentratsioonigradienti. Gradient säilib, kui plasmas lahustunud hapnik on seotud hemoglobiiniga, s.t. hapniku difusioon veest toimub seni, kuni hemoglobiin on täielikult hapnikuga küllastunud. Vere hapnikumaht ulatub astelrail 65 mg/l kuni lõhe 180 mg/l. Vere küllastumine süsihappegaasiga (süsinikdioksiid) võib aga vähendada kalade vere hapnikumahtu 2 korda.
Süsinikdioksiid transporditakse veres erinevalt. Hemoglobiini roll süsinikdioksiidi ülekandmisel karbohemoglobiini kujul on väike. Arvutused näitavad, et hemoglobiin ei kanna rohkem kui 15% kalade ainevahetuse tulemusena tekkivast süsihappegaasist. Süsinikdioksiidi ülekande peamine transpordisüsteem on vereplasma.
Rakkudest difusiooni tulemusena verre sattudes tekitab süsinikdioksiid oma piiratud lahustuvuse tõttu plasmas suurenenud osarõhu ja peaks seega pärssima gaasi üleminekut rakkudest vereringesse. Tegelikkuses seda ei juhtu. Plasmas toimub reaktsioon erütrotsüütide karboanhüdraasi toimel
Tänu sellele väheneb pidevalt süsihappegaasi osarõhk rakumembraanil vereplasma küljelt ning süsihappegaasi difusioon verre toimub ühtlaselt. Karboanhüdraasi roll on skemaatiliselt näidatud joonisel fig. 8.4.
Saadud bikarbonaat siseneb koos verega lõpuseepiteeli, mis sisaldab ka karboanhüdraasi. Seetõttu muutuvad vesinikkarbonaadid lõpustes süsihappegaasiks ja veeks. Lisaks difundeerub CO 2 kontsentratsioonigradienti mööda verest vette, pestes lõpuseid.
Lõpusefilamentide kaudu voolav vesi puutub lõpuseepiteeliga kokku mitte rohkem kui 1 sekundiks, mistõttu süsihappegaasi kontsentratsioonigradient ei muutu ja see väljub vereringest ühtlase kiirusega. Süsinikdioksiid eemaldatakse ligikaudu samal viisil teistes hingamisteedes. Lisaks eritub organismist märkimisväärses koguses ainevahetuse tulemusena tekkivat süsihappegaasi karbonaatidena uriiniga, kõhunäärmemahlaga, sapiga ja naha kaudu.
Testtund teemal “Kalad”
Kui palju huvitavat olete kalade kohta varasemates tundides ja lisakirjandusest teada saanud! Kas saate vastata järgmistele küsimustele?
1. Miks on elus kala käes raske hoida? (Soomuste väliskülg on kaetud limakihiga, mida eritavad nahanäärmed. Lima vähendab hõõrdumist kala keha ja vee vahel ning kaitseb bakterite ja hallituse eest.)
2. Miks ei jookse kala isegi mudases vees takistustele? (Kaladel on spetsiaalne meeleorgan – külgjoon.)
3. Miks haid ei uppu, kuigi neil pole ujupõit? (Hai keha ujuvus saavutatakse tänu suurte rasvavarude kuhjumisele eelkõige maksas. Seetõttu ulatub mõnel hailiigil maksa mass 25%-ni kogu kehamassist, luukaladel aga on vaid 1–8%.)
4. Miks mõned kalaliigid munevad palju mune? (Neid ei iseloomusta järglaste eest hoolitsemine; nad hülgavad oma munad "saatuse meelevalda" - enamiku munadest ja maimudest söövad röövloomad.)
5. Milline kala on selles neljas veider (vt pilti)? (Hai on kõhrekalade klassi esindaja.)
6. Kellel on pikem seedesüsteem: haugil või hõbekarpkal? (Hõbekarpkalal; soolestiku pikkus sõltub toidu iseloomust: röövkaladel on see palju lühem kui taimtoidulistel kaladel.)
7. Kui palju vereringet on kaladel? (Üks, välja arvatud kopsukalad – neil on kopsud.)
8. Mis on see ajuosa (mudel näitab väikeaju) ja miks see kaladel nii suur on? (Väikeaju. See kontrollib looma liigutuste koordineerimist ja tasakaalu, mis on eriti oluline veekeskkonnas.)
9. Millised elundid peale lõpuste võivad kalade hingamises osaleda? (Ujumispõis, kopsud (kopskaladel), sooled, nahk (kui kala kehal puuduvad soomused), epibranhiaalne labürint.)
Lisaküsimused
1. Nüüd võib kirjandusteostest leida bioloogilisi vigu. Näiteks kolm autorit, mainides sama looma, teevad vigu:
A.K. Tolstoi eeposes “Sadko”: “Ja siin/Beluga vaatab teda uudishimulikult, silmi pilgutades...”
Sasha Cherny: "Ja mu kallis naine / ohkab nagu beluga."
Boriss Pasternak ühineb nendega filmis "Doktor Živago": "Jaamades mürisesid auruvedurid nagu beluga..."
(Beluuga on kala ja loomulikult ta ei pilguta – kaladel pole silmalaugusid. Beluga ei "möirga" ega "ohka"; see erineb täiesti erinevast loomast, beluga vaalast, imetajast, polaardelfiin.)
2. Kuid mitte kõik kalad pole vaiksed. Mõned neist võivad teha erinevaid helisid. Selles aitab neid sageli orel, mis võib samuti tugevdada tajutavaid helisid. Mis organ see on, milliseid funktsioone see veel täidab?
(Ujumispõis on hüdrostaatiline aparaat, vere gaaside sisalduse regulaator ja mõnel liigil täiendav hingamiselund.)
3. Tooge näide toiduahelast, mis hõlmab meie piirkonnas leiduvaid kalaliike.
(Antud näites peab olema vähemalt kaks kalaliiki.)