Белок состоящий только из остатков аминокислот. Пространственная организация белковых молекул
Белки – это биологические полимеры, состоящие из аминокислот. Ни один из существующих живых организмов – от вирусов до растений и животных – не может существовать без белка. Правда, у растений имеются особые возбудители болезней – вироиды, состоящие из одной нуклеиновой кислоты, однако для их размножения необходимы белки растительной клетки-хозяина.
Белки выполняют в организме множество жизненно важных функций.
Структурная функция
Структурную функцию выполняет, например, белок кератин, из которого состоят шерсть, рога, копыта, верхний отмерший слой кожи. В зависимости от числа поперечных сшивок, скрепляющих белковые молекулы, кератиновые структуры бывают довольно мягкими и гибкими (волосы), а бывают чрезвычайно жесткими и прочными (панцирь черепахи).
В сухожилиях содержится белок коллаген, его фибриллы почти не поддаются растяжению. Благодаря этому мышечное усилие передается костям, к которым крепятся мышцы. При кипячении в воде коллаген образует желатину, часто применяющуюся для приготовления студней и желе. Белок эластин, наоборот, не слишком прочен, но очень эластичен, он содержится в стенках сосудов, легко растягивающихся при увеличении давления.
Белки выполняют структурную функцию не только на организменном, но и на клеточном уровне – в любой эукариотической клетке есть состоящий из белков внутренний цитоскелет. Различают три различных цитоскелетных системы: микротрубочки, микрофиламенты и промежуточные филаменты.
Микротрубочки представляют собой трубчатые образования, состоящие из белка тубулина. По ним, как по рельсам, движутся органеллы от одного участка клетки к другому (другие белки прикрепляют органеллы к наружной стороне «трубы» и обеспечивают движение). Во время митоза они обеспечивают расхождение хромосом к полюсам клетки.

Рис. 1. Слева – строение микротрубочки, справа – цитоскелет, образованный микротрубочками, в клетке соединительной ткани – фибробласте. Микротрубочки окрашены зеленым, ядро – голубым
Микрофиламенты состоят из белка актина. Они образуют сплошную сеть под наружной мембраной клетки, придавая ей упругость и прочность. Пучки микрофиламентов образуются на переднем конце движущейся амебы (и любой клетки с амебоидным движением), именно они выпячивают ложноножку (псевдоподию).

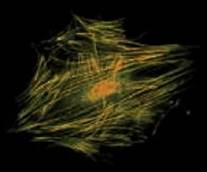
Рис. 2. Слева – строение микрофиламента, справа – цитоскелет, образованный микрофиламентами, в фибробласте. Микрофиламенты окрашены желтым
Промежуточные филаменты в разных клетках состоят из различных белков. В эпителиальных клетках они состоят из кератина, так что волосы представляют собой остатки мертвых ороговевших клеток. По-видимому, эти филаменты просто придают механическую прочность клетке.
Каталитическая функция
Катализатор – это вещество, которое ускоряет реакцию, оставаясь в конце ее неизменным (не расходуясь). Биологические катализаторы называются ферментами, а вещества, участвующие в самой реакции, – субстратами. Почти все ферменты – это белки. В живой клетке может содержаться около 1000 ферментов.
Для живой клетки весьма ценны такие особенности работы ферментов по сравнению с обычными «химическими» катализаторами, как специфичность, высокая эффективность и регулируемость.
Обычно один фермент узнает только «свой» субстрат и ускоряет одну определенную реакцию. Правда, в некоторых случаях специфичность нужна лишь в определенных пределах – так, многие протеазы расщепляют любую пептидную связь белкового субстрата, они неспецифичны к аминокислотным остаткам, составляющим эту связь. Однако они не расщепляют связи между остатками моносахаридов.
Большинство ферментов значительно превосходят по каталитической активности неорганические и простые органические катализаторы. Для эффективной работы небиологических катализаторов, как правило, нужна высокая температура, тогда как в организме человека все ферменты обходятся температурой около 37°С (а у холоднокровных животных – и более низкой).
Еще одно ценное свойство ферментов – это регулируемость, т.е. способность «включаться» и «выключаться». Это относится не ко всем ферментам, некоторые и не надо регулировать.
Однако у ферментов есть и недостатки. Так, они не выдерживают высокой температуры – теряют свою каталитическую активность из-за денатурации (впрочем, у разных белков различная устойчивость к температурным воздействиям – у бактерий-термофилов белки нормально работают при 100 °С).
Многие ферменты нуждаются для своей работы в наличии небольших небелковых соединений – коферментов. Они часто образуются из витаминов – почти все витамины группы В являются предшественниками коферментов. Некоторые коферменты прочно связаны со своими ферментами, тогда как другие легко отделяются от одного белка и присоединяются к другому.
Некоторые ферменты активны только тогда, когда связываются с ионами металлов – магния, марганца, цинка, железа, меди и др.
Все известные способы движения живых организмов основаны на работе соответствующих белков. Так, сокращение мышц обеспечивают мышечные белки актин и миозин. В поперечно-полосатых мышцах имеются пучки актиновых и миозиновых нитей, которые называются тонкими и толстыми филаментами. При возбуждении мышцы эти филаменты начинают скользить друг по другу. Толстые филаменты как бы втягиваются в пространство между тонкими, в результате чего мышца сокращается (энергию для такого направленного скольжения дает АТФ).
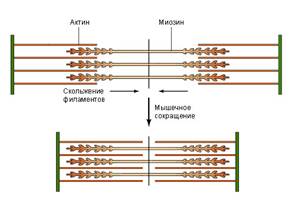
Рис. 3. Скольжение актиновых и миозиновых нитей вызывает мышечное сокращение
Они же делают возможным ползание амебы. На переднем конце амебоидной клетки растут актиновые филаменты, они выпячивают наружную мембрану, образуя ложноножку. Затем ложноножка прикрепляется к поверхности, по которой ползет амеба. Наконец, с помощью миозина вся клетка подтягивается к прикрепленной ложноножке, и процесс повторяется снова.
Другие белки обеспечивают подвижность жгутиков. По окружности жгутика эукариотических клеток располагаются микротрубочки, связанные друг с другом с помощью белка динеина. Этот белок как бы пытается заставить скользить одну микротрубочку по другой (вспомните скольжение нитей при мышечном сокращении). Но микротрубочки скреплены друг с другом специальными белками, поэтому они не могут свободно скользить друг по другу, а могут лишь изгибаться. Этот изгиб распространяется по всему жгутику, он начинает биться как хлыст, вызывая движение всей клетки. Динеин работает на энергии АТФ.
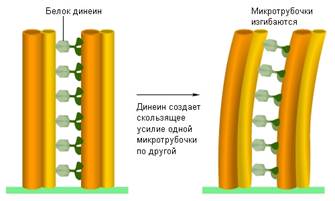
Рис. 4. Механизм движения жгутика эукариотических клеток
Транспортная функция
Классический пример транспортного белка – это гемоглобин крови, который переносит кислород по кровяному руслу (он участвует и в транспорте углекислого газа). Имеются специальные белки, переносящие по организму различные вещества: ионы железа (белок трансферрин), витамин В12 (транскобаламин), жирные кислоты (сывороточный альбумин), стероидные гормоны и т. п.
Специальные белки служат и для транспорта разных веществ через мембрану. Глюкоза является гидрофильным соединением и очень плохо проникает через липидный бислой, поэтому на мембранах различных клеток имеется специальный белок – переносчик глюкозы. Хотя бислой проницаем для воды, все же на мембране есть белки–аквапорины, ускоряющие прохождение воды через нее. Некоторые аквапорины специфично транспортируют только воду, другие могут переносить еще и разные небольшие нейтральные молекулы (глицерин, мочевину).
Питательная или энергетическая функция
Белки можно расщепить, окислить и получить энергию, необходимую для жизни. При окислении 1 г белка выделяется около 4,1 килокалории. Обычно белки идут на энергетические нужды организма человека в крайних случаях, когда исчерпаны запасы жиров и углеводов.
В яйцеклетках содержатся специальные запасные белки (например, яичный альбумин). Когда начинается развитие нового организма из оплодотворенного яйца, они расщепляются и используются как «строительный материал» для синтеза новых белков, а также как источник энергии. Запасные белки содержатся и в семенах растений.
Защитная функция
В крови и других жидкостях содержатся белки, которые могут убивать или помогать обезвреживать микробов. В состав плазмы крови входят антитела – белки, каждый из которых узнает определенный вид микроорганизмов или иных чужеродных агентов, – а также защитные белки системы комплемента. Существует несколько классов антител (эти белки еще называют иммуноглобулинами), самый распространенный из них – иммуноглобулин G. В слюне и в слезах содержится белок лизоцим – фермент, расщепляющий муреин и разрушающий клеточные стенки бактерий. При заражении вирусом клетки животных выделяют белок интерферон, препятствующий размножению вируса и образованию новых вирусных частиц.
Защитную функцию для микроорганизмов выполняют и такие неприятные для нас белки, как микробные токсины – холерный токсин, токсин ботулизма, дифтерийный токсин и т. п. Повреждая клетки нашего организма, они защищают микробов от нас.
Рецепторная функция
Белки служат для восприятия и передачи сигналов. В физиологии есть понятие клетки-рецептора, т.е. клетки, которая воспринимает определенный сигнал (например, в сетчатке глаза находятся клетки-зрительные рецепторы). Но в клетках-рецепторах эту работу осуществляют белки–рецепторы. Так, белок родопсин, содержащийся в сетчатке глаза, улавливает кванты света, после чего в клетках сетчатки начинается каскад событий, который приводит к возникновению нервного импульса и передаче сигнала в мозг.
Белки-рецепторы есть не только в клетках-рецепторах, но и в других клетках. Очень важную роль в организме играют гормоны – вещества, выделяемые одними клетками и регулирующие функцию других клеток. Гормоны связываются со специальными белками – рецепторами гормонов на поверхности или внутри клеток-мишеней.
Многие (хотя и далеко не все) гормоны являются белками – например, все гормоны гипофиза и гипоталамуса, инсулин и др. Еще одним примером белков, выполняющих эту функцию, могут служить внутриклеточные белки, регулирующие работу генов.
Многие белки могут выполнять несколько функций.
Макромолекулы белков состоят из α-аминокислот. Если в состав полисахаридов обычно входит одна и та же «единица» (иногда две), повторяющаяся много раз, то белки синтезируются из 20 разных аминокислот. После того, как молекула белка собрана, некоторые аминокислотные остатки в составе белка могут подвергаться химическим изменениям, так что в «зрелых» белках можно обнаружить более 30 различных аминокислотных остатков. Такое разнообразие мономеров обеспечивает и многообразие биологических функций, выполняемых белками.
α-аминокислоты имеют следующее строение:

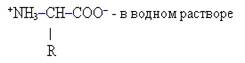
здесь R – различные группы атомов (радикалы) у разных аминокислот. Ближайший к карбоксильной группе атом углерода обозначается греческой буквой α, именно с этим атомом соединена аминогруппа в молекулах α-аминокислот.
В нейтральной среде аминогруппа проявляет слабые основные свойства и присоединяет ион Н+, а карбоксильная – слабо кислотные и диссоциирует с освобождением этого иона, так что хотя в целом суммарный заряд молекулы не изменится, она будет одновременно нести положительно и отрицательно заряженную группу.
В зависимости от природы радикала R различают гидрофобные (неполярные), гидрофильные (полярные), кислые и щелочные аминокислоты.
У кислых аминокислот имеется вторая карбоксильная группа. Она немного сильнее карбоксильной группы уксусной кислоты: у аспарагиновой кислоты половина карбоксилов диссоциирована при рН 3,86, у глютаминовой – при рН 4,25, а у уксусной – лишь при 4,8. Среди щелочных аминокислот самой сильной является аргинин: половина его боковых радикалов сохраняет положительный заряд при рН 11,5. У лизина боковой радикал является типичным первичным амином, он остается наполовину ионизированным при рН 9,4. Самая слабая из щелочных аминокислот – гистидин, его имидазольное кольцо наполовину протонировано при рН 6.
Среди гидрофильных (полярных) также имеются две аминокислоты, способные ионизироваться при физиологических рН – цистеин, у которого SH-группа может отдавать ион Н+ подобно сероводороду, и тирозин, у которого есть слабокислая фенольная группировка. Однако эта способность выражена у них очень слабо: при рН 7 цистеин ионизирован на 8 %, а тирозин – на 0,01 %.
Для обнаружения α-аминокислот обычно используют нингидриновую реакцию: при взаимодействии аминокислоты с нингидрином образуется ярко окрашенный синий продукт. Кроме того, отдельные аминокислоты дают свои специфические качественные реакции. Так, ароматические аминокислоты дают желтое окрашивание с азотной кислотой (в ходе реакции происходит нитрование ароматического кольца). При подщелачивании среды окраска изменяется на оранжевую (подобное изменение окраски происходит и у индикаторов, например, метилоранжа). Эта реакция под названием ксантопротеиновой используется также для детекции белка, поскольку в большинстве белков есть ароматические аминокислоты; желатин не дает этой реакции, поскольку почти не содержит ни тирозина, ни фенилаланина, ни триптофана. При нагревании с плюмбитом натрия Na2PbO2 цистеин образует черный осадок сульфида свинца PbS.
Растения и многие микробы могут синтезировать аминокислоты из простых неорганических веществ. Животные могут синтезировать лишь некоторые аминокислоты, другие же должны получать с пищей. Такие аминокислоты называются незаменимыми. Для человека незаменимыми являются фенилаланин, триптофан, треонин, метионин, лизин, лейцин, изолейцин, гистидин, валин и аргинин. К сожалению, злаковые культуры содержат очень мало лизина и триптофана, зато эти аминокислоты в существенно большем количестве содержатся в бобовых культурах. Не случайно традиционные диеты земледельческих народов обычно содержат как злаки, так и бобовые: пшеница (или рожь) и горох, рис и соя, кукуруза и бобы являются классическими примерами такого сочетания у народов разных континентов.
α-Атом углерода у всех 20 аминокислот находится в состоянии sp3-гибридизации. Все его 4 связи расположены под углом около 109°, так что формулу аминокислоты можно вписать в тетраэдр.
Легко убедиться, что могут существовать два вида аминокислот, которые являются зеркальными отображениями друг друга. Как бы мы ни перемещали и ни поворачивали их в пространстве, совместить их невозможно – они различаются как правая и левая рука.
Такой вид изомерии называется оптической изомерией. Он возможен только в том случае, если у центрального атома углерода (он называется асимметрическим центром) со всех 4 сторон находятся разные группы (поэтому глицин не имеет оптических изомеров, а остальные 19 аминокислот имеют). Из двух разных изомерных форм аминокислот ту, что на рис. 1 расположена справа, называют D-формой, а слева – L-формой.
Основные физические и химические свойства D- и L-изомеров аминокислот одинаковы, однако различаются оптические свойства: их растворы вращают плоскость поляризации света в противоположные стороны. Различна и скорость их реакций с другими оптически активными соединениями.
Интересно, что в состав белков всех живых организмов от вирусов до человека входят только L-аминокислоты. D-формы встречаются в некоторых антибиотиках, синтезируемых грибами и бактериями. Белки могут образовывать упорядоченную структуру лишь в том случае, если в их состав будут входить только изомеры аминокислот одного типа.
Строение белковых молекул
В молекуле белка аминокислотные остатки соединены так называемой пептидной связью. Полная последовательность аминокислотных остатков в такой цепи называется первичной структурой белка. Число остатков в разных белках может колебаться от нескольких штук до нескольких тысяч. Небольшие молекулы с мол. массой менее 10 тыс. дальтон называют пептидами, а крупные – белками. В составе белка обычно имеются как кислые, так и щелочные аминокислоты, так что белковая молекула имеет и положительные, и отрицательные заряды. Значение рН, при котором количество отрицательных зарядов равно количеству положительных, называется изоэлектрической точкой белка.
Обычно белковая цепочка складывается в более сложные структуры. Кислород группы C=O может образовывать водородную связь с водородом группы N–H, расположенной в другой аминокислоте. За счет таких водородных связей формируется вторичная структура белка. Одна из разновидностей вторичной структуры – α-спираль. В ней каждый кислород С=О-группы связан с водородом 4-й по ходу спирали NH-группы. На один виток спирали приходится 3,6 аминокислотных остатка, шаг спирали составляет 0,54 нм.
Во многих белках имеется т. н. β-структура, или β-слой, в ней полипептидные цепочки почти полностью развернуты, их отдельные участки своими группами –СО– и –NH– образуют водородные связи с другими участками той же цепочки или соседней полипептидной цепи.
α-Спиральную структуру имеет белок кератин, из которого состоят волосы и шерсть. При нагревании влажные волосы и шерсть легко поддаются растяжению, а потом самопроизвольно возвращаются к исходному состоянию: при растяжении водородные связи α-спирали разрываются, а затем постепенно восстанавливаются.
β-Структура характерна для фиброина – основного белка шелка, выделяемого гусеницами шелкопряда. В отличие от шерсти, шелк почти нерастяжим – β-структура образована вытянутыми полипептидными цепями, и дальше растягивать ее практически невозможно без разрыва ковалентных связей.
Укладка белков обычно не ограничивается вторичной структурой. Гидрофобные аминокислотные остатки «стремятся» укрыться от водного окружения внутри белковой молекулы. Между боковыми группами кислых и щелочных аминокислот, заряженных, соответственно, отрицательно и положительно, возможно электростатическое взаимодействие. Многие аминокислотные остатки могут образовывать друг с другом водородные связи. Наконец, остатки аминокислоты цистеина, содержащие SH-группы, способны образовывать между собой ковалентные связи –S–S–.
Благодаря всем этим взаимодействиям – гидрофобным, ионным, водородным и дисульфидным – белковая цепочка образует сложную пространственную конфигурацию, называемую третичной структурой.
В составе глобулы у многих белков можно различить отдельные компактные участки размером около 10–20 тысяч дальтон. Они называются доменами. Участки полипептидной цепи между доменами весьма гибки, так что всю структуру можно представить как относительно жесткие бусины доменов, соединенные гибкими промежуточными участками первичной структуры.
Многие белки (их называют олигомерными) состоят не из одной, а из нескольких полипептидных цепочек. Совокупность их образует четвертичную структуру белка, при этом отдельные цепочки называются субъединицами. Четвертичная структура удерживается теми же связями, что и третичная. Пространственная конфигурация белка (т.е. его третичная и четвертичная структура) называется конформацией.
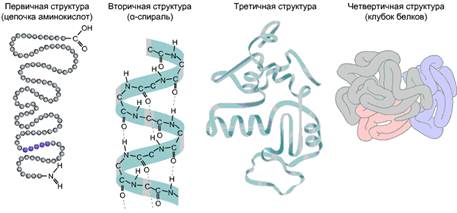
Рис. 4. Различные уровни укладки белковых молекул
Главным методом, позволяющим устанавливать пространственную структуру белков и других биологических полимеров, является рентгеноструктурный анализ. В последнее время большие успехи достигнуты в компьютерном моделировании белковых конформаций.
Водородные, электростатические и гидрофобные связи, с помощью которых создаются вторичная, третичная и четвертичная структуры белка, менее прочны, чем пептидная связь, образующая первичную структуру. При нагревании они легко разрушаются, и хотя у белка сохраняется в целости первичная структура, он не может выполнять своих биологических функций, становится неактивным. Процесс разрушения природной конформации белка, сопровождающийся потерей активности, называется денатурацией. Денатурацию вызывает не только нагревание, но и химические вещества, нарушающие связи вторичной и третичной структур – например, мочевина, которая в высоких концентрациях разрушает водородные связи в белковой глобуле.
Дисульфидные –S–S-связи образуют прочные «скрепы», сшивающие различные участки одной полипептидной цепи или разные цепи. Эти связи имеются, например, в кератинах, причем разные кератины содержат разное количество таких сшивок: волосы и шерсть – немного, рога, копыта млекопитающих и панцири черепах – значительно больше.
Вторичная, третичная и четвертичная структура белка определяется его первичной структурой. В зависимости от последовательности аминокислот в полипептидной цепочке будут образовываться α-спиральные или β-структурные участки, которые затем самопроизвольно «уложатся» в определенную третичную структуру, а у некоторых белков отдельные цепочки еще и объединятся с образованием четвертичной структуры.
Если изменить первичную структуру белка, то может сильно измениться и вся его конформация. Существует тяжелая наследственная болезнь – серповидно-клеточная анемия, при которой гемоглобин становится малорастворим в воде, а эритроциты приобретают серповидную форму. Причина болезни – замена всего лишь одной аминокислоты из 574, входящих в состав человеческого гемоглобина (глютаминовая кислота, расположенная на 6-м месте с N-конца одной из цепей гемоглобина нормальных людей, у больных заменена на валин).
Процесс самопроизвольной ассоциации белковых субъединиц в сложные комплексы, обладающие четвертичной структурой, называется самосборкой. Большинство белковых комплексов с четвертичной структурой образуются именно путем самосборки.
В 1980-е годы было обнаружено, что не все белки и белковые комплексы образуются путем самосборки. Оказалось, что для образования таких структур как нуклеосомы (комплексы белков-гистонов с ДНК), бактериальные ворсинки – пили, а также некоторых сложных ферментных комплексов используются специальные белки–помощники, названные шаперонами. Шапероны не входят в состав образующейся структуры, а только помогают ее укладке.
Шапероны служат не только для организации сложных комплексов, но в некоторых случаях помогают правильно свернуть одну полипептидную цепочку. Так, при воздействии высокой температуры в клетках резко возрастает количество т. н. белков теплового шока. Они связываются с частично денатурированными клеточными белками и восстанавливают их природную конформацию.
Долгое время считалось, что белок может иметь только одну устойчивую конформацию при данных условиях, но в последнее время этот постулат пришлось пересмотреть. Поводом к такому переосмыслению послужило открытие возбудителей т. н. медленных нейрологических инфекций. Эти инфекции встречаются у разных видов млекопитающих. К ним относится болезнь овец «скрепи», заболевание человека «куру» («смеющаяся смерть») и нашумевшее в последнее время «бешенство коров». Они имеют много общего.
Для них характерны тяжелые поражения центральной нервной системы. Так, люди, больные куру, на ранних стадиях болезни испытывают эмоциональную неустойчивость (большинство – часто и беспричинно смеются, но некоторые находятся в состоянии депрессии или немотивированной агрессивности) и легкие нарушения координации движений. На поздних стадиях больные уже не способны не только двигаться, но даже сидеть без поддержки, а также есть.
Заражение обычно происходит пищевым путем (изредка через кровь). Болезнь у животных развивалась после скармливания им костной муки, которая изготавливалась из костей больных особей. Куру – болезнь папуасских людоедов, передающаяся при поедании мозга умерших родичей (поедание друг друга в этом случае – скорее отрасль богослужения, чем кулинарии, оно имеет важное ритуальное значение).
Все эти заболевания имеют очень длительный инкубационный период и развиваются медленно. В головном мозге заболевших отмечается отложение нерастворимого белкового конгломерата. Нерастворимые белковые нити обнаруживаются в пузырьках, расположенных внутри нейронов, а также во внеклеточном веществе. Наблюдается разрушение нейронов в некоторых отделах мозга, особенно в мозжечке.
Долгое время оставалась загадочной природа возбудителей этих заболеваний, и только в начале 80-х годов было установлено, что эти возбудители – особые белки с молекулярной массой около 30 тысяч дальтон. Такие доселе неизвестные науке объекты получили название прионы.
Было установлено, что прионный белок закодирован в ДНК организма-«хозяина». Белок здорового организма содержит ту же последовательность аминокислот, что и белок инфекционной частицы приона, но никаких патологических симптомов не вызывает. Функция прионного белка пока неизвестна. Мыши, у которых генные инженеры искусственно выключили ген этого белка, развивались вполне нормально, хотя и имели некоторые отклонения в работе центральной нервной системы (худшее обучение, нарушения сна). В здоровом организме этот белок обнаружен на поверхности клеток во многих органах, больше всего в головном мозге.
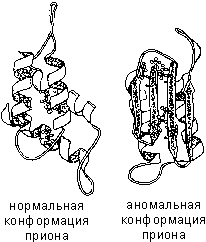
Оказалось, что в инфекционной частице прионный белок имеет иную конформацию, чем в нормальных клетках. Он содержит бета-структурные участки, обладает высокой устойчивостью к расщеплению пищеварительными ферментами и способностью образовывать нерастворимые агрегаты (видимо, отложение таких агрегатов в мозге и является причиной развития нейропатологии).
Самое интересное, что «нормальная» конформация этого белка превращается в «болезнетворную», если клетка контактирует с «болезнетворным» белком. Получается, что «болезнетворный» белок «лепит» пространственную структуру «нормального» по себе. Он направляет его укладку как матрица, вызывая появление все большего числа молекул в «болезнетворной» конформации и, в конце концов, гибель организма.
Как именно это происходит, до сих пор неизвестно. Если смешать в пробирке нормальную и инфекционную форму прионного белка, то никакого образования новых инфекционных молекул не произойдет. По-видимому, в живой клетке имеются какие-то молекулы-помощники (вероятно, шапероны), позволяющие прионному белку делать свое черное дело.
Отложение нерастворимых белковых конгломератов может служить причиной и других неизлечимых нервных заболеваний. Болезнь Альцгеймера не относится к числу инфекционных – она возникает в пожилом и старческом возрасте у людей с наследственной предрасположенностью. У больных наблюдается ухудшение памяти, ослабление интеллекта, слабоумие, и, в конце концов – полная утрата психических функций. Причина развития болезни – отложение в мозгу т. н. амилоидных бляшек. Они состоят из нерастворимого белка – β-амилоида. Он представляет собой фрагмент белка–предшественника амилоида – нормального белка, присутствующего у всех здоровых людей. У больных он расщепляется с образованием нерастворимого амилоидного пептида.
Мутации в разных генах вызывают развитие болезни Альцгеймера. Естественно, ее вызывают мутации в гене белка–предшественника амилоида – измененный предшественник после расщепления образует нерастворимый β-амилоид, формирующий бляшки и разрушающий клетки головного мозга. Но болезнь возникает и при мутации в генах белков, регулирующих активность протеаз, разрезающих белок – предшественник амилоида. Не совсем понятно, как в этом случае развивается заболевание: возможно, нормальный белок предшественник режется в каком-то неправильном месте, что приводит к выпадению в осадок образующегося пептида.
Очень рано болезнь Альцгеймера развивается у больных с синдромом Дауна – у них содержится не две копии 21-й хромосомы, как у всех людей, а три. Больные синдромом Дауна имеют характерную внешность и слабоумие. Дело в том, что ген белка–предшественника амилоида как раз и находится в 21-й хромосоме, увеличение количества гена приводит к увеличению количества белка, а избыток белка–предшественника приводит к накоплению нерастворимого β-амилоида.
Белки часто соединяются с другими молекулами. Так, гемоглобин, переносящий кислород в кровеносной системе, состоит из белковой части – глобина, и небелковой – гема. Ион Fe2+ входит в состав гема. Глобин состоит из четырех полипептидных цепочек. Благодаря наличию гема с железом гемоглобин катализирует окисление перекисью водорода различных органических веществ – например, бензидина. Раньше эта реакция под названием «бензидиновая проба» использовалась в судебно-медицинской экспертизе для обнаружения следов крови.
Некоторые белки химически связаны с углеводами, их называют гликопротеины. Очень многие белки, секретируемые животной клеткой, являются гликопротеинами – например, известные по предыдущим разделам трансферрин и иммуноглобулины. Однако, желатин, хотя и является продуктом гидролиза секретируемого белка коллагена, практически не содержит присоединенных углеводов. Внутри клетки гликопротеины встречаются гораздо реже.
В лабораторной практике используется много методов определения концентрации белка. В самом простом из них применяется биуретовый реактив – щелочной раствор соли двухвалентной меди. В щелочной среде некоторая часть пептидных связей в молекуле белка переходит в енольную форму, которая образует с двухвалентной медью комплексы, окрашенные в красный цвет. Другой распространенной реакцией на белок является окраска по Бредфорду. В ходе реакции молекулы специального красителя связываются с белковой глобулой, что вызывает резкое изменение окраски – из бледно-коричневого раствор становится ярко-синим. Это краситель – «кумасси ярко-синий» – раньше использовался для окраски шерсти (а шерсть, как известно, состоит из белка кератина). Наконец, для определения концентрации белка можно использовать его свойство поглощать ультрафиолетовый свет с длиной волны 280 нм (его поглощают ароматические аминокислоты фенилаланин, тирозин и триптофан). Чем сильнее раствор поглощает такой ультрафиолет, тем больше там содержится белка.
Щелочная ср. NH2 R R R COOH COO – COO – Катион Амфион Анион Таким образом, фактором, определяющим поведение белка как катиона или аниона, является реакция среды, которая определяется концентрацией водородных ионов и выражается величиной рН. Однако...
Строение белков
Белки - высокомолекулярные органические соединения, состоящие из остатков α-аминокислот.
В состав белков входят углерод, водород, азот, кислород, сера. Часть белков образует комплексы с другими молекулами, содержащими фосфор, железо, цинк и медь.
Белки обладают большой молекулярной массой: яичный альбумин - 36 000, гемоглобин - 152 000, миозин - 500 000. Для сравнения: молекулярная масса спирта - 46, уксусной кислоты - 60, бензола - 78.
Аминокислотный состав белков
Белки - непериодические полимеры, мономерами которых являются α-аминокислоты . Обычно в качестве мономеров белков называют 20 видов α-аминокислот, хотя в клетках и тканях их обнаружено свыше 170.
В зависимости от того, могут ли аминокислоты синтезироваться в организме человека и других животных, различают: заменимые аминокислоты - могут синтезироваться; незаменимые аминокислоты - не могут синтезироваться. Незаменимые аминокислоты должны поступать в организм вместе с пищей. Растения синтезируют все виды аминокислот.
В зависимости от аминокислотного состава, белки бывают: полноценными - содержат весь набор аминокислот; неполноценными - какие-то аминокислоты в их составе отсутствуют. Если белки состоят только из аминокислот, их называют простыми . Если белки содержат помимо аминокислот еще и неаминокислотный компонент (простетическую группу), их называют сложными . Простетическая группа может быть представлена металлами (металлопротеины), углеводами (гликопротеины), липидами (липопротеины), нуклеиновыми кислотами (нуклеопротеины).
Все аминокислоты содержат : 1) карбоксильную группу (–СООН), 2) аминогруппу (–NH 2), 3) радикал или R-группу (остальная часть молекулы). Строение радикала у разных видов аминокислот - различное. В зависимости от количества аминогрупп и карбоксильных групп, входящих в состав аминокислот, различают: нейтральные аминокислоты , имеющие одну карбоксильную группу и одну аминогруппу; основные аминокислоты , имеющие более одной аминогруппы; кислые аминокислоты , имеющие более одной карбоксильной группы.
Аминокислоты являются амфотерными соединениями , так как в растворе они могут выступать как в роли кислот, так и оснований. В водных растворах аминокислоты существуют в разных ионных формах.
Пептидная связь
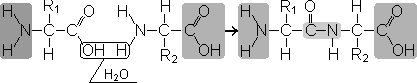
Пептиды - органические вещества, состоящие из остатков аминокислот, соединенных пептидной связью.
Образование пептидов происходит в результате реакции конденсации аминокислот. При взаимодействии аминогруппы одной аминокислоты с карбоксильной группой другой между ними возникает ковалентная азот-углеродная связь, которую и называютпептидной . В зависимости от количества аминокислотных остатков, входящих в состав пептида, различают дипептиды, трипептиды, тетрапептиды и т.д. Образование пептидной связи может повторяться многократно. Это приводит к образованиюполипептидов . На одном конце пептида находится свободная аминогруппа (его называют N-концом), а на другом - свободная карбоксильная группа (его называют С-концом).
Пространственная организация белковых молекул
Выполнение белками определенных специфических функций зависит от пространственной конфигурации их молекул, кроме того, клетке энергетически невыгодно держать белки в развернутой форме, в виде цепочки, поэтому полипептидные цепи подвергаются укладке, приобретая определенную трехмерную структуру, или конформацию. Выделяют 4 уровня пространственной организации белков .
Первичная структура белка - последовательность расположения аминокислотных остатков в полипептидной цепи, составляющей молекулу белка. Связь между аминокислотами - пептидная.

Если молекула белка состоит всего из 10 аминокислотных остатков, то число теоретически возможных вариантов белковых молекул, отличающихся порядком чередования аминокислот, - 10 20 . Имея 20 аминокислот, можно составить из них еще большее количество разнообразных комбинаций. В организме человека обнаружено порядка десяти тысяч различных белков, которые отличаются как друг от друга, так и от белков других организмов.
Именно первичная структура белковой молекулы определяет свойства молекул белка и ее пространственную конфигурацию. Замена всего лишь одной аминокислоты на другую в полипептидной цепочке приводит к изменению свойств и функций белка. Например, замена в β-субъединице гемоглобина шестой глутаминовой аминокислоты на валин приводит к тому, что молекула гемоглобина в целом не может выполнять свою основную функцию - транспорт кислорода; в таких случаях у человека развивается заболевание - серповидноклеточная анемия.
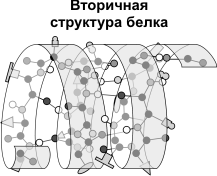
Вторичная структура - упорядоченное свертывание полипептидной цепи в спираль (имеет вид растянутой пружины). Витки спирали укрепляются водородными связями, возникающими между карбоксильными группами и аминогруппами. Практически все СО- и NН-группы принимают участие в образовании водородных связей. Они слабее пептидных, но, повторяясь многократно, придают данной конфигурации устойчивость и жесткость. На уровне вторичной структуры существуют белки: фиброин (шелк, паутина), кератин (волосы, ногти), коллаген (сухожилия).
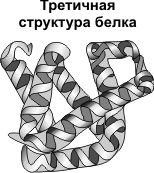

Третичная структура - укладка полипептидных цепей в глобулы, возникающая в результате возникновения химических связей (водородных, ионных, дисульфидных) и установления гидрофобных взаимодействий между радикалами аминокислотных остатков. Основную роль в образовании третичной структуры играют гидрофильно-гидрофобные взаимодействия. В водных растворах гидрофобные радикалы стремятся спрятаться от воды, группируясь внутри глобулы, в то время как гидрофильные радикалы в результате гидратации (взаимодействия с диполями воды) стремятся оказаться на поверхности молекулы. У некоторых белков третичная структура стабилизируется дисульфидными ковалентными связями, возникающими между атомами серы двух остатков цистеина. На уровне третичной структуры существуют ферменты, антитела, некоторые гормоны.
Четвертичная структура характерна для сложных белков, молекулы которых образованы двумя и более глобулами. Субъединицы удерживаются в молекуле благодаря ионным, гидрофобным и электростатическим взаимодействиям. Иногда при образовании четвертичной структуры между субъединицами возникают дисульфидные связи. Наиболее изученным белком, имеющим четвертичную структуру, является гемоглобин . Он образован двумя α-субъединицами (141 аминокислотный остаток) и двумя β-субъединицами (146 аминокислотных остатков). С каждой субъединицей связана молекула гема, содержащая железо.
Если по каким-либо причинам пространственная конформация белков отклоняется от нормальной, белок не может выполнять свои функции. Например, причиной «коровьего бешенства» (губкообразной энцефалопатии) является аномальная конформация прионов - поверхностных белков нервных клеток.
Свойства белков
Аминокислотный состав, структура белковой молекулы определяют его свойства . Белки сочетают в себе основные и кислотные свойства, определяемые радикалами аминокислот: чем больше кислых аминокислот в белке, тем ярче выражены его кислотные свойства. Способность отдавать и присоединять Н + определяют буферные свойства белков ; один из самых мощных буферов - гемоглобин в эритроцитах, поддерживающий рН крови на постоянном уровне. Есть белки растворимые (фибриноген), есть нерастворимые, выполняющие механические функции (фиброин, кератин, коллаген). Есть белки активные в химическом отношении (ферменты), есть химически неактивные, устойчивые к воздействию различных условий внешней среды и крайне неустойчивые.
Внешние факторы (нагревание, ультрафиолетовое излучение, тяжелые металлы и их соли, изменения рН, радиация, обезвоживание)
могут вызывать нарушение структурной организации молекулы белка. Процесс утраты трехмерной конформации, присущей данной молекуле белка, называют денатурацией . Причиной денатурации является разрыв связей, стабилизирующих определенную структуру белка. Первоначально рвутся наиболее слабые связи, а при ужесточении условий и более сильные. Поэтому сначала утрачивается четвертичная, затем третичная и вторичная структуры. Изменение пространственной конфигурации приводит к изменению свойств белка и, как следствие, делает невозможным выполнение белком свойственных ему биологических функций. Если денатурация не сопровождается разрушением первичной структуры, то она может быть обратимой , в этом случае происходит самовосстановление свойственной белку конформации. Такой денатурации подвергаются, например, рецепторные белки мембраны. Процесс восстановления структуры белка после денатурации называется ренатурацией . Если восстановление пространственной конфигурации белка невозможно, то денатурация называется необратимой .
Функции белков
| Функция | Примеры и пояснения |
| Строительная | Белки участвуют в образовании клеточных и внеклеточных структур: входят в состав клеточных мембран (липопротеины, гликопротеины), волос (кератин), сухожилий (коллаген) и т.д. |
| Транспортная | Белок крови гемоглобин присоединяет кислород и транспортирует его от легких ко всем тканям и органам, а от них в легкие переносит углекислый газ; в состав клеточных мембран входят особые белки, которые обеспечивают активный и строго избирательный перенос некоторых веществ и ионов из клетки во внешнюю среду и обратно. |
| Регуляторная | Гормоны белковой природы принимают участие в регуляции процессов обмена веществ. Например, гормон инсулин регулирует уровень глюкозы в крови, способствует синтезу гликогена, увеличивает образование жиров из углеводов. |
| Защитная | В ответ на проникновение в организм чужеродных белков или микроорганизмов (антигенов) образуются особые белки - антитела, способные связывать и обезвреживать их. Фибрин, образующийся из фибриногена, способствует остановке кровотечений. |
| Двигательная | Сократительные белки актин и миозин обеспечивают сокращение мышц у многоклеточных животных. |
| Сигнальная | В поверхностную мембрану клетки встроены молекулы белков, способных изменять свою третичную структуру в ответ на действие факторов внешней среды, таким образом осуществляя прием сигналов из внешней среды и передачу команд в клетку. |
| Запасающая | В организме животных белки, как правило, не запасаются, исключение: альбумин яиц, казеин молока. Но благодаря белкам в организме могут откладываться про запас некоторые вещества, например, при распаде гемоглобина железо не выводится из организма, а сохраняется, образуя комплекс с белком ферритином. |
| Энергетическая | При распаде 1 г белка до конечных продуктов выделяется 17,6 кДж. Сначала белки распадаются до аминокислот, а затем до конечных продуктов - воды, углекислого газа и аммиака. Однако в качестве источника энергии белки используются только тогда, когда другие источники (углеводы и жиры) израсходованы. |
| Каталитическая | Одна из важнейших функций белков. Обеспечивается белками - ферментами, которые ускоряют биохимические реакции, происходящие в клетках. Например, рибулезобифосфаткарбоксилаза катализирует фиксацию СО 2 при фотосинтезе. |
Ферменты
Ферменты , или энзимы , - особый класс белков, являющихся биологическими катализаторами. Благодаря ферментам биохимические реакции протекают с огромной скоростью. Скорость ферментативных реакций в десятки тысяч раз (а иногда и в миллионы) выше скорости реакций, идущих с участием неорганических катализаторов. Вещество, на которое оказывает свое действие фермент, называют субстратом .
Ферменты - глобулярные белки, по особенностям строения ферменты можно разделить на две группы: простые и сложные.Простые ферменты являются простыми белками, т.е. состоят только из аминокислот. Сложные ферменты являются сложными белками, т.е. в их состав помимо белковой части входит группа небелковой природы - кофактор . У некоторых ферментов в качестве кофакторов выступают витамины. В молекуле фермента выделяют особую часть, называемую активным центром.
Активный центр - небольшой участок фермента (от трех до двенадцати аминокислотных остатков), где и происходит связывание субстрата или субстратов с образованием фермент-субстратного комплекса. По завершении реакции фермент-субстратный комплекс распадается на фермент и продукт (продукты) реакции. Некоторые ферменты имеют (кроме активного) аллостерические центры - участки, к которым присоединяются регуляторы скорости работы фермента (аллостерические ферменты ).
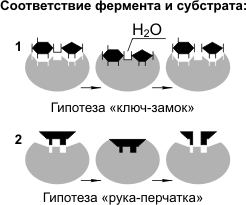
Для реакций ферментативного катализа характерны: 1) высокая эффективность, 2) строгая избирательность и направленность действия, 3) субстратная специфичность, 4) тонкая и точная регуляция. Субстратную и реакционную специфичность реакций ферментативного катализа объясняют гипотезы Э. Фишера (1890 г.) и Д. Кошланда (1959 г.).
Э. Фишер (гипотеза «ключ-замок») предположил, что пространственные конфигурации активного центра фермента и субстрата должны точно соответствовать друг другу. Субстрат сравнивается с «ключом», фермент - с «замком».
Д. Кошланд (гипотеза «рука-перчатка») предположил, что пространственное соответствие структуры субстрата и активного центра фермента создается лишь в момент их взаимодействия друг с другом. Эту гипотезу еще называют гипотезой индуцированного соответствия .
Скорость ферментативных реакций зависит от: 1) температуры, 2) концентрации фермента, 3) концентрации субстрата, 4) рН. Следует подчеркнуть, что поскольку ферменты являются белками, то их активность наиболее высока при физиологически нормальных условиях.
Большинство ферментов может работать только при температуре от 0 до 40 °С. В этих пределах скорость реакции повышается примерно в 2 раза при повышении температуры на каждые 10 °С. При температуре выше 40 °С белок подвергается денатурации и активность фермента падает. При температуре, близкой к точке замерзания, ферменты инактивируются.
Билет 1.
Цепи,связывают,остатков,связями.
Белки,взаимодействий,структуру,конформацией.
Последовательность,содержит,о построении,трехмерной.
Уровня различают,называемых.
Линейная,остатков,в цепи,пер белка.
Индивидуального каждого,в участке ДНК,геном.
В,синтеза,информ-ия в гене перп,на м-РНК,рибосоме,сборка пер струк.
Каждый из 50 тыс,белков,имеет уникальную.
Все моле-лы, белка,одинаковое,остатков,в белке,отличает,другого.
Цепи,связывают,связями.
Взаимодействий,сруктуру,конформацией.
Последовательность,третичную.
Последовательность.
Закодирована,ДНК,геном.
Синтеза,мРНК,сборка.
Каждый,уникальную.
Первичная структура белка.
В настоящее время расшифрована первичная структура около 2500 белков, а в природе имеется 10 12 разнообразных белков.
Первичная структура – это последовательность (порядок) соединения аминокислотных остатков с помощью пептидной связи.
Пептидная связь образуется за счет карбоксильной группы одной аминокислоты и аминогруппы другой аминокислоты.
В образовании первичной структуры участвуют -аминокислоты.
Пептидная связь образует остов полипептидной цепи, она является повторяющимся фрагментом.
Свойства первичной структуры белка
Детерминированность – последовательность аминокислот в белке генетически закодирована. Информация о последовательности аминокислот содержится в ДНК.
Уникальность – для каждого белка в организме характерна определенная последовательность аминокислот.
Аминокислоты, входящие в состав белков делят на 2 группы:
Взаимозаменяемые аминокислоты – это амиокислоты, сходные по структуре и свойствам.
Невзаимозаменяемые аминокислоты, отличающиеся по структуре и свойствам.
В белковой молекуле различают 2 вида замен аминокислот:
Консервативная – замена одной аминокислоты на другую сходную по структуре. Такая замена не приводит к изменению свойств белка.
Примеры: гли-ала, асп-глу, тир-фен, вал-лей.
Радикальная замена – замена одной аминокислоты на другую отличающуюся по структуре. Такая замена приводит к изменению свойств белка.
Примеры: глу-вал, сер-цис, про-три, фен-асп, илей-мет.
При радикальной замене возникает белок с другими свойствами, что может привести к патологии.
3.Универсальность первичной структуры. Белки, выполняющие одинаковые функции в разных организмах имеют одинаковую или близкую первичную структуру.
В природных белках одна и та же аминокислота не встречается подряд больше 3 раз.
Радикальная замена глу на вал в шестом положении в молекуле гемоглобина приводит к развитию серповидно-клеточной анемии. При этой патологии эритроциты в условиях низкого парциального давления приобретают форму серпа. После отдачи кислорода такой гемоглобин превращается в плохо растворимую форму и начинает выпадать в осадок в виде веретенообразных кристаллоидов,. Тактоиды деформируют клетку и эритроциты приобретают форму серпа. При этом происходит гемолиз эритроцитов. Болезни протекает остро и дети погибают. Эта патология называется серповидно-клеточной анемией.
Классическим примером наследственной гемоглобинопатии является серповидноклеточная анемия.Глу в 6-м положении в бетта –цепи заменен на валин.Эритроциты в условиях низкого парциального давления кислорода принимают форму серпа.Такой гемоглобин после отдачи кислорода превращается в плохо растворимую форму и начинает выпадать в осадок в виде веретенообразных кристаллоидов, названных тактоидами,которые деформируют клетку и вызывают массивных гемолиз.
Любой дефект рецептра ЛПНП или белка апоВ-100,взаимодействующего с ним,приводит к развитию наиболее распространенного наследственного заболевания –семейной гиперхолестеринемии.Пичиной этого аутосомно-доминантного заболевания выступают мутации в гене рецептора ЛПНП.Гетерозиготы,имеющие один нор ген,встречаются с част-й 1:500.Количество рецепторов наа поверхности клеток у них снижено вдвое,а конц холестерола в плазме повышается вдвое.Это приводит к выраженному атеросклерозу и инфаркту или инсульту.Гомозиготы-1:1млн.У них конц хол увеличена в 5-6 раз.
Связывая ЛНП, клетки поддерживают определенный уровень этих ЛП в крови. У всех обследованных здоровых людей интернализация ЛНП неизбежно сопровождается и связыванием с рецепторами клеток. Связывание и интернализация ЛНП обеспечивается одним и тем же белком, входящим в состав рецепторов ЛНП. В фибробластах больных с семейной гиперхолестеринемией, дефицитных по рецепторам ЛНП интернализация их резко угнетается.
Появление в клетках свободного холестерина ингибирует ОМГ-КоА-редуктазу снижает эндогенный синтез холестерина. При концентрации ЛНП > 50 мкг/мл синтез холестерина в фибробластах подавляется полностью.
Гиперхолестеринемия – главная причина отложения холестерина в артериях. Но важное значение имеют также первичные повреждения стенок сосудов. Повреждения эндотелия могут возникать в следствие гипертонии, воспалительных процессов.
В области повреждения эндотелия в стенку сосудов проникают компоненты крови, в том числе липопротеиды, которые поглощаются макрофагами. Мышечные клетки сосудов начинают размножаться и тоже фагоцитировать липопротеиды. Ферменты лизосом разрушают липопротеиды, кроме холестерина. Холестерин накапливается в клетке, клетка гибнет, а холестерин оказывается в межклеточном пространстве и инкапсулируется соединительной тканью – образуется атеросклеротическая бляшка.
Между отложением холестерина в артериях и липопротеидами крови происходит обмен, но при гиперхолестеринемии преобладает поток холестерина в стенки сосудов.
Методы профилактики и лечения атеросклероза направлены на уменьшение гиперхолестеринемии. Для этого применяют малохолестериновую диету, лекарства увеличивающие эксткрецию холестерина или ингибирующие его синтез, прямое удаление холестерина из крови методом гемодиффузии.Печень донора с нормальным количеством рецепторов.Резкое повышение холестерина в пище (до 1,5 г ежедневно) может сопровождаться некоторой гиперхолестеринемией у здоровых людей.
Стр455 Северин.
Билет 2.
Конформация – это пространственное расположение в органической молекуле замещающих групп, способных свободно изменять свое положение в пространстве без разрыва связей, благодаря свободному вращению вокруг одинарных углеродных связей.
Различают 2 вида вторичной структуры белка:
1.
 -спираль
-спираль
2.
 -складчатость.
-складчатость.
Вторичную структуру стабилизируют водородные связи. Водородные связи возникают между атомом водорода в NH группе и карбоксильным кислородом.
Характеристика
-спирали.
Для
каждого белка характерна своя степень
спирализации полипептидной цепи.
Спирализованные участки чередуются с
линейными. В молекуле гемоглобина
и-цепи
спирализованы на 75%, в лизоциме – 42%,
пепсине – 30%.
Степень спирализации зависит от первичной структуры белка.
пролин.его атом азота входит в состав жесткого кольца,что исключает возможность вращения вокруг N-CH связей.Кроме того,у атома азота пролина,образующего связь сдругой аминокислотой нет водорода.В результате пролин не способен организовать водородную связь и структура альфа спиралей нарушается.Обычно здесь возникает петля или изгиб.
Участки,где последовательно расположены несколько одинаково заряженных радикалов,между которыми возникают электростатические силы отталкивания.
Участки с близко расположенными обьемными радикалами,механически нарушающими формирование альфа спиралей,например метионин,триптофан.
Спирализации белковой молекулы препятствует аминокислота пролин.
-
складчатость имеет слабоизогнутую
конфигурацию полипептидной цепи.
Если
связанные полипептидныые цепи направлены
противоположно,возникает антипараллельная
-структура,если
жеN
и С концы полипептидных цепей
совпадают,возникает структура
параллельного
-складчатого
слоя.
Для
-
складчатости характерны водородные
связи в пределах одной полипептидной
цепи или сложных полипептидных цепей.
В
белках возможны переходы от
-спирали
к-складчатости
и обратно вследствие перестройки
водородных связей.
-складчатость
имеет плоскую форму.
-спираль
имеет стержневую форму.
Водородные связи – слабые связи, энергия связи 10 – 20 ккал/моль, но большое количество связей обеспечивает стабильность белковой молекулы.
Билет 3.
Третичная структура белка.
Третичной структурой белка называется трехмерная простанственная структура образующаяся за счет взаимодействия между радикалами аминокислот,которые могут располагаться на знаачительном расстоянии в цепи.
По форме третичной структуры белка делят на глобулярные и фибриллярные.
В стабилизации третичной структуры белковой молекулы участвуют ковалентные связи (пептидные и дисульфидные). Основную роль в стабилизации играют нековалентные связи: водородные, электростатические взаимодействия заряженных групп, межмолекулярные ван-дер-вальсовы силы, взаимодействия неполярных боковых радикалов аминокислот, так называемые гидрофобные взаимодействия.
Электростатические=ионные могут возникать внутри гидрофоб ядра между анионными ггруппами аспартата и глутамата и катионными лизина,аргинина,гистидина.
Гидрофобные радикалы аминокислот ала, вал, изолей, мет, фен в водной среде взаимодействуют друг с другом. При этом неполярные гидрофобные радикалы аминокислот как бы погружаются внутрь белковой молекулы, образуя там сухие зоны, а полярные радикалы оказываются ориентированными в сторону воды.В результате внутри белковой глобулы оказывается гидрофобное ядро.
Если же белки функционируют в липидном окружении,то наоборот. В любом случае радикалы аминокислот занимают наиболее выгодное биоэнергетическое положение.
При укладке полипептидная цепь белка стремится принять энергетически выгодную форму с меньшим запахом энергии.
При формировании третичной структуры полипептидная цепь изгибается в местах нахождения пролина, глицина.
Глобулярные белки растворимы в воде, а фибриллярные нет.
Активные центры формируются за счет ограниченного числа аминокислот (12-16). Часто аминокислоты удалены друг от друга. Активные центры возникают при образовании четвертичной структуры.
Сближение различных фунх грп,прин\х раз\м уч-м п ц\и позвол\т соз\ть простра\ю конф\ю контактного уч\а акт\го ц\ра.Т е форма а ц закладывается,а формируется при чет или при добии коф\та.
Билет 4
Денатурация.
Денатурация – это процесс нарушения высших уровней организации белковой молекулы (вторичного, третичного, четвертичного) под действием различных факторов.
При этом полипептидная цепь разворачивается и находится в растворе в развернутом виде или в виде беспорядочного клубка.
При денатурации утрачивается гидратная оболочка и белок выпадает в осадок и при этом утрачивает нативные свойства.
Денатурацию вызывают физические факторы: температура, давление, механические воздействия, ультразвуковые и ионизирующие излучения; химические факторы: кислоты, щелочи, органические растворители, алкалоиды, соли тяжелых металлов.
Различают 2 вида денатурации:
Обратимая денатурация – ренатурация или ренактивация – это процесс, при котором денатурированный белок, после удаления денатурирующих веществ вновь самоорганизуется в исходную структуру с восстановлением биологической активности.
необратимая денатурация – это процесс, при котором биологическая активность не восстанавливается после удаления денатурирующих агентов.
Свойства денатурированных белков.
Увеличение числа реактивных или функциональных групп по сравнению с нативной молекулой белка (это группы COOH, NH 2 , SH, OH, группы боковых радикалов аминокислот).
Уменьшение растворимости и осаждение белка (связано с потерей гидратной оболочки), развертыванием молекулы белка, с «обнаружением» гидрофобных радикалов и нейтрализации зарядов полярных групп.
Изменение конфигурации молекулы белка.
Потеря биологической активности, вызванная нарушением нативной структуры.
Более легкое расщепление протеолитическими ферментами по сравнению с нативным белком – переход компактной нативной структуры в развернутую рыхлую форму облегчает доступ ферментов к пептидным связям белка, которые они разрушают.
Ферментные методы гидролиза основаны на избирательности действия протеолитических ферментов расщепляющих пептидные связи между определенными аминокислотами.
Пепсин расщепляет связи, образованные остатками фенилаланина, тирозина и глутаминовой кислоты.
Трипсин расщепляет связи между аргинином и лизином.
Химотрипсин гидролизует связи триптофана, тирозина и фенилаланина.
Гидрофобные взаимодействия,а также ионные и водородные связи относятся к числу слабых,тк энергия их лишь ненамного превосходит энергию теплового движения атомов при комнатной температуре(т е уже при данной температуре возможен разрыв связей).
Поддержание характерной для белка конформации возможно благодаря возникновению множества слабых связей между различными участками полипептидной цепи.
Однако,белки состоят из огромного числа атомов,находящихся в посттояном (броуновском) движении,что приводит к енбольшим перемещениям отдельных участков полипептидной цепи,которые обычно не нарушают общую структуру белка и его функции.Следовательно,белки обладают конформационной лабильностью – склонностью к небольшим изменениям конформации за счет разрыва одних и образования других слабых связей.Конформация белка может меняться при изменении химических и физических средств среды,а также при взаимодействии белка с другими молекулами.При этом происходит изменение пространственной структуры не только участка,контактирующего с другой молекулой,но и конформации белка в целом.Конформационные изменения играют роль огромную в функционировании белков в клетке живой.
Билет 5
Четвертичная структура белка-это количество и взаиморасположение полипептидных цепей
Белки, состоящие из одной полипептидной цепи, имеют только третичную структуру (лизоцим, пепсин, миоглобин, трипсин).Их называют мономерами.Цепи белков соединенные ковалентными связями (например дисульфидными)поэтому инсулин мономерный белок.
Для белков, состоящих из нескольких полипептидных цепей, характерна четвертичная структура.
Под четвертичной структурой понимают объединение отдельных полипептидных цепей с третичной структурой в функционально активную молекулу белка. Каждая отдельная полипептидная цепь называется протомером и чаще не обладает биологической активностью.Олигомерные белки содержат от 2(гексокиназа)до 312(пируватдегидрогеназа)пртомеров.Специфичность связывания протомеров за счет зависит от совокупности радикалов третичной структуры и определяется комплементарностью протомеров.
Комплементарность-пространственное и химическое соответствие взаимодействующих поверхностей.
В молекуле белка может быть несколько протомеров, которые при объединении образуют олигомер или мультимер.
Для белков с четвертичной структурой характерно понятие субъединицы.
Субъединица – это функционально активная часть молекулы белка.
Примером
белка с четвертичной структурой является
гемоглобин, состоящий из 4 протомеров:
2
и 2-цепей.
Взаимодействие полипептидных цепей при формировании олигомера происходит за счет полярных групп аминокислотных остатков. Между полярными группами образуется ионная, водородные связи, гидрофобные взаимодействия.
Активные центры возникают при образовании четвертичной структуры.
В молекуле белка имеются прочные (ковалентные) связи, а также слабые, что обеспечивает с одной стороны стабильность молекулы, а с другой лабильность.
Альфа спирали в протомере обозначают латинскими буквами от A до H,начиная с N конца
Кооперативные изменения конформации протомеров.
Кислород связывается с протомерами гемоглобина чере железо(2),который соединен с 4 атомами азота пиррольных колец и атомом азота Гис F8 белковой части протомера.Связывание кислорода с оставшейся координационной связью железа происходит по другую сторону от плоскости гема.Гис Е7 обеспечивает оптимальные условия.Присоединение кислорода к атому железа одного протомера вызывает его перемещение в плоскостьь гема,за ним перемещаются остаток Гис F8 и полипептидная цепь.Так как протомер связан с остальными протомерами,а белки обладают конформационной лабильностью,происходит изменение конформации всего белка.Конформационные изменения,произошедшие в других протомерах,облегчают присоединение следующей молекулы кислорода,что вызывает новые конформационные изменения в белке и ускорение связывания следующей молекулы кислорода.Четвертая молекула кислорода присоединяется к гемоглобину в 300 раз легче первой.
Изменение конформации(а следовательно и функциональных свойств) всех протомеров олигомерного белка при присоединение лиганда только к одному из них носит название Кооперативные изменения конформации протомеров.
Аналогичным образом в тканях диссоцифция каждой молекулы кислорода изменяет конфоормацию всех протомеров и облегчает отщепление последующих молекул кислорода.
Билет 6
Классификация белков.
По хим строению.
простые белки-это те,которые содержат в своем составе только полипептидные цепи,состоящие из аминокислотных остатков.Пример:основные белкки хроматина-гистоны;эластин.
сложные белки-те,которые,кроме полипептидной цепи,содержат в своем составе небелковую часть,присоединенную слабыми или ковалентными связями.Небелковая часть может быть представлена ионами металлов,какими-либо органическими молекулами.Прочно связанная с белком небелковая часть носит название простетической группы:гемопротеины,цитохромы,каталаза,пероксидаза,фосфопротеины,липопротеины,гллликопротеины,Сложный белок,т.о.,состоит из небелковой части(прстетическая группа,и белковой(апопротеин) и называется холопротеином.
По функциям.
ферменты-это специализированные белки,ускоряющие течение химических реакций.Благодаря ф. скорости хим реакций возрастают в миллиооны раз.Благодаря набору ферментов в клетках превращения поступающих вещесттвв протекают не хаотично,а в строго определенных направлениях.
регуляторрные белки-это большая группа белковых гормонов,участвующих в поддержании постоянства внутренней среды организма.Они воздействуют на клетки мишени.Например:инсулин,кальмодулиин(присоед 4 кальция,крепится к др б\м и рег\т их фун\ю)
рецепторные белки-для гидрофобных молекул они внутри клетки.
транспортные –переносят молекулы,плохо растворимые в воде: альбумин переноситв плазме жир к\ты и билирубин,гемоглобин.
Структурные белки-они расположены определенным образом в тканях,придают им форму,создают опору,определяют механические свойства данной ткани:коллаген,эластин.
защитные белки-иммуноглобулины,фибриноген,протромбин.
Сократительные белки-наделяют клетку способностью либо сокращаться,либо передвигаться.:актин,миозин,тубулин,
Физико-химические свойства белков:это различие индивидуальных белков по форме молекул,по массе,суммарному заряду,соотношению полярных и неполярных групп,растворимость,степень устойчивости к воздействию денатурирующих агентов.
по форме молекул на глобуллярные и фибриллярные.
У глобулярных компактна структтура,гидрофобные радикалы спрятаны в гидрофобное ядро,лучше растворимы в жидкостях организма.
По молекулярной массе:белки-вмс,номогут сильно оттличаться по молекулярной массе,а именно: от 6000 1млн Д в зависимости от коол\ва ам\т и протомеров.
Суммарный заряд белков: они имеют в своем составе радикалы лизина,аргинина,гистидина,глутамата и аспартата,способные к ионизации.Степень ионизации зависит от рН среды.При 7 все ионогенныегр б нах в ионниз состоянии.В кислой среде происходит подавление иониз глу и асп,в щелочеой –уменьш «+» заряда. Изоэлектрическая точка-значение рН,при котором белок приобретает суммарный нулевой заряд. Тк в большенстве белков преобладают глу и асп,то рI для большинства белков лежит в слабокислой среде..гистоны-для них в слабощелочной.Белки,имеющие «-« или «+» заряд лучше растворимы.Суммарный заряд ув кол-во диполей воды,способных связаться с белком, и препятствует контакту одноименнно заряженных молекул,в результате чего повышает растворимость в воде.Заряженные белки могут двигаться в электрическом поле.
Соотношение полярных и неполярных групп на поверхности нативных молекул белков.Протомеры олигомерныхбелков часто в области контактов содержат гидрофобные радикалы.индивидуальлно для индив\х белков.
Растворимост белков-зависит от всех вышеперечисленных свойств белков:формы,м м\ы,величины заряда,соотношения полярных и неполярных групп на пов б\а.Кроме того,р\ть опред\ся составом растворителя,напр,некоторые белки легче растворяются в слабом солевом ра\ре,нежели в дистиллированной воде.
Билет 7.
Структура и свойства ферментов.
Ферменты (энзимы) – специфические белки, входящие в состав всех клеток и тканей живых организмов, играющие роль биологических катализаторов.
Доказательства белковой природы ферментов.
Изоэлектрическая точка ферментов.
Поведение ферментов при изменении концентрации водородных генов.
Высокая специфичность ферментов.
Ферменты не способны проникать через полупроницаемые мембраны.
Сохранение активности ферментами после действия водоотнимающими средствами (ацетон, спирт, нейтральные соли щелочных металлов).
Инативация ферментов при нагревании. Инактивация ферментов совпадает с денатурацией белка. Ферменты разрушаются также под действием минеральных кислот, щелочей, солей, алкалоидов, при облучении рентгеновскими и ультрафиолетовыми лучами.
Электрохимические свойства ферментов.
Различают следующие виды активных центров:
Субстратный (якорная площадка) активный центр – обеспечивает присоединение субстрата за счет образования слабых связей: водородных, ван-дер-ваальсовых, гидрофобных взаимодействий.
Каталитический активный центр – отвечает за превращение субстрата. В пространстве эти центры могут быть разделены, а могут быть совмещены.
Аллостерический (регуляторный) обеспечивает присоединение низкомолекулярных веществ, приводит к изменению активности фермента. Аллостерический центр удален от субстратного и каталитического центров.
Закономерности построения активных центров.
Активные центры формируются за счет ограниченного числа аминокислот (12-16). Часто аминокислоты удалены друг от друга. Активные центры возникают при образовании четвертичной структуры.
В построении активных центров часто участвуют аминокислоты: гис, сер, лиз, асп, цис.
В построении активных центров сложных ферментов участвуют группировки кофакторов.
Олиго- и мультимерные ферменты на каждом протомере имеют свой каталитический и субстратный центр, аллостерический центр формируется за счет нескольких протомеров. При разрушении четвертичной структуры нарушается аллостерический центр и регуляция прекращается, а каталитическая функция характерная для протомера сохраняется.
Активный центр – это трехмерная структура, имеющая вид впадины или щели.
Теории, объясняющие механизм взаимодействия фермента и субстрата.
Теория Фишера – теория предшествующего соответствия, теория «ключ – замок». Согласно теории активный центр фермента существует и точно соответствует субстрату.
Недостатки (противоречия) теории:
Нет соответствия в термодинамических расчетах (разница в расчетном количестве выделяемой энергии и практически выделяемом количестве энергии).
По этой теории фермент может ошибаться и присоединять похожий субстрат.
Субстраты часто низкомолекулярные вещества, а ферменты высокомолекулярные, содержащие большое число аминокислот. Теория не объясняла существование групповой специфичности.
Теория Кошленда – индуцированного соответствия, т.е. активный центр формируется в момент взаимодействия фермента и субстрата, т.е. происходит подгонка. В субстрате происходит изменение связей. Наличие активных центров определяют специфичность.
Виды специфичности.
Абсолбтная – одному субстрату соответствует один фермент.
Уреаза катализирует расщепление мочевины, аспартаза катализирует взаимодействие NH 3 с фумаровой кислотой, в результате образуется аспарагиновая кислота, но аспартаза не действует на малеиновую кислоту.
|| 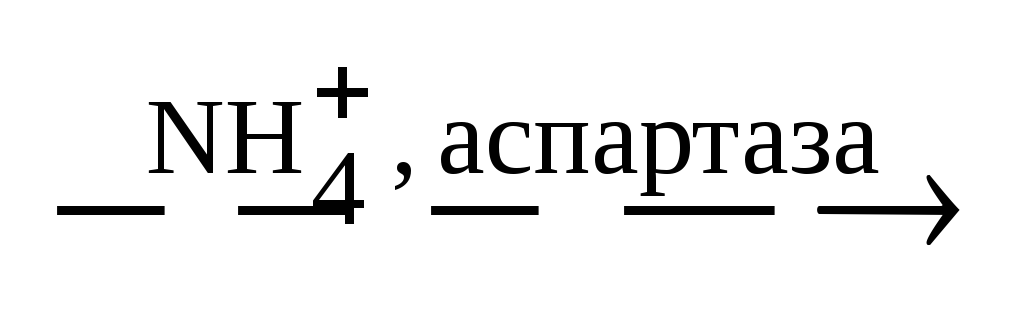 |
|
CH 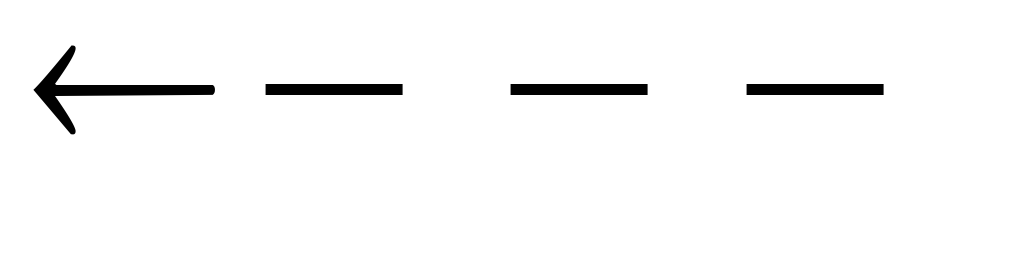 CHNH 2
CHNH 2
Фумарат Аспартат
Относительная групповая специфичность. Фермент расщепляет группу субстратов, для которых характерен один тип связей.
Пример: пепсин расщепляет пептидную связь –CO-NH между аминокислотами.
Аналогично действуют трипсин, химотрипсин, пептидазы.
Химотрепсин расщепляет пептидные связи между три, тир и фен, но при определенных условиях могут расщеплять амидные и сложноэфирные связи.
Стереохимическая специфичность – фермент обеспечивает превращение определенного оптического (стереоизомера). В организме происходит превращение L-аминокислот, но D-углеводов.
Фумараза катализирует превращение фумаровой кислоты (транс-изомер), но не действует на малеиновую кислоту (цис-изомер).
При исследовании специфичности ферментов было установленно, что молекула субстрата должна обладать двумя структурными особенностями:
Субстрат должен содержать специфическую химическую связь, которую фермент мог атаковать.
В молекуле субстрата должна быть функциональная группа, называемая связывающей группой, которая способна связываться с ферментом и ориентировать молекулу субстрата в активном центре фермента, чтобы атакуемая связь субстрата была правильно расположена по отношению к каталитической группе фермента.
БИЛЕТ №8.
Концентрация субстрата зависит от питания, возраста, физической нагрузки.
Зависимость скорости ферментативной реакции субстрата выражается уравнением Михаэлиса-Ментен:
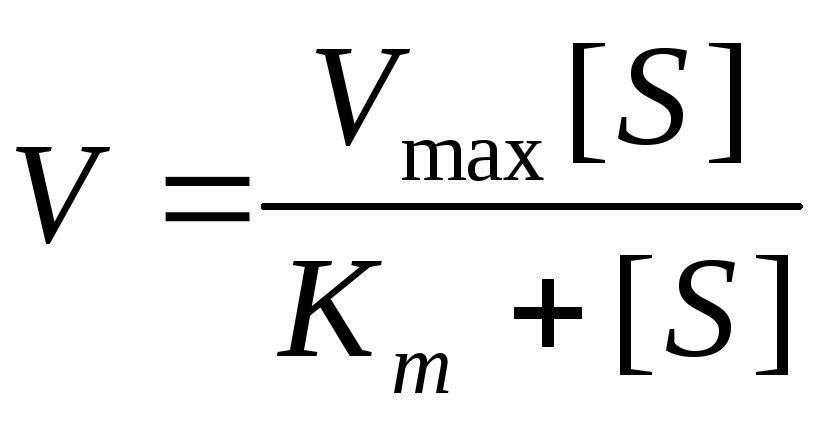
V max – максимальная скорость реакции
[S] – концентрация субстрата
K m – константа Михаэлиса.
Анализ уравнения Михаэлиса-Ментен.
Концентрация субстрата мала, стремиться к нулю. При этих условиях [S] можно пренебречь:
[S] 0,
при этом [S]
можно пренебречь:
0,
при этом [S]
можно пренебречь:
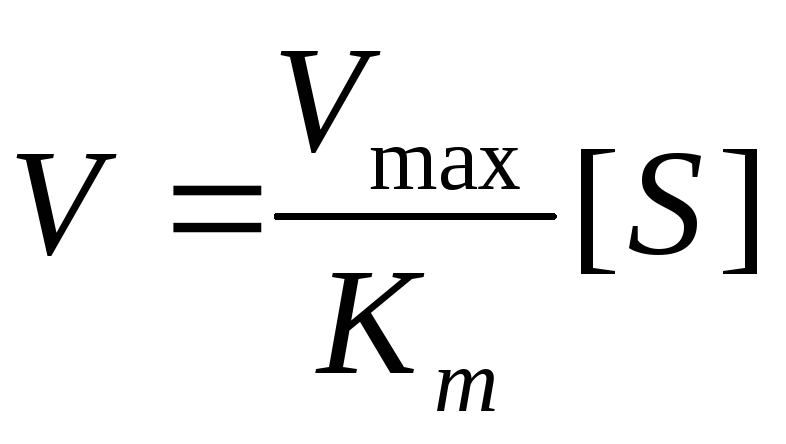
Концентрация субстрата стремится к бесконечности, пренебрегаем K m и уравнение имеет вид:
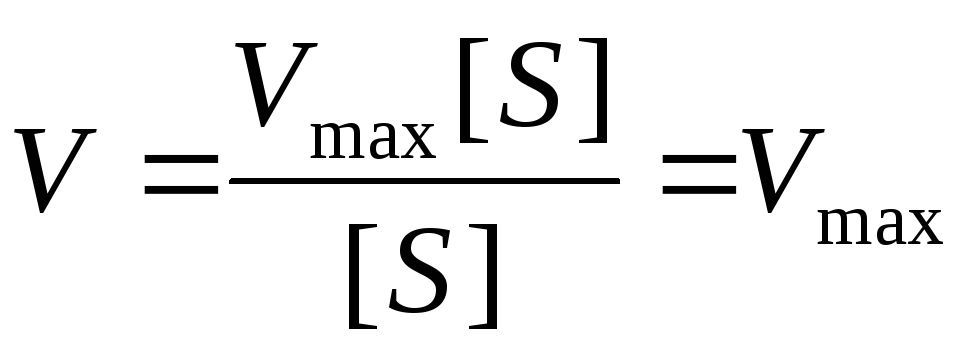
Сокращаем на [S] и скорость реакции равняется V max .
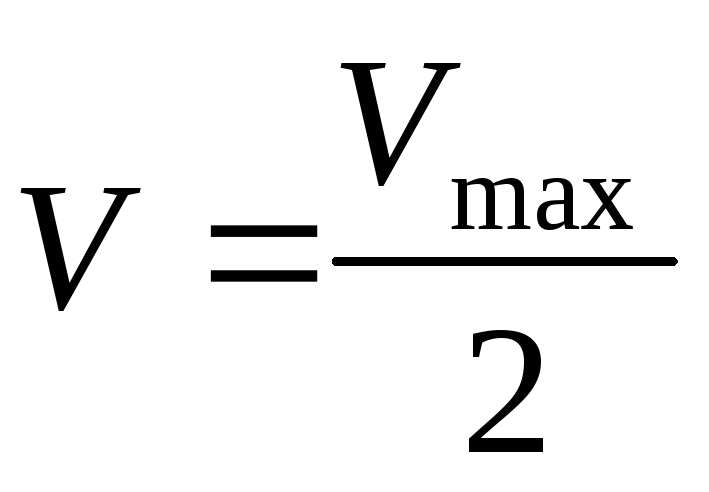
 и разделив на
V max
получим
и разделив на
V max
получим
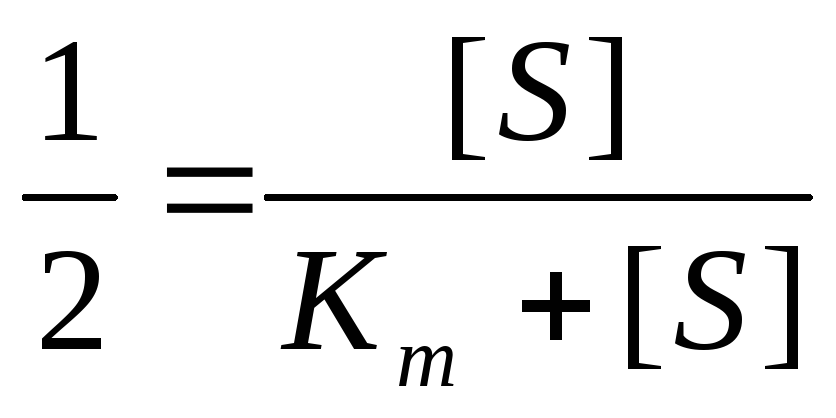 .
Решая уравнение относительноK m
получаем K m +[S]
= 2[S],
.
Решая уравнение относительноK m
получаем K m +[S]
= 2[S],
K m
– величина, численно равная концентрации
субстрата при
,
выраженная в молях.K m
= 10 -1
– 10 -6
– для клеток организма, величина const.
K m показывает:
Степень сродства между ферментом и субстратом, существует обратная зависимость – чем меньше K m , тем больше сродство F к S.
K m позволяет определить какой субстрат будет превращаться под действием данного фермента:
Например, этиленгликоль – составная часть антифриза и алкагольдегидрогеназа (АДГ) будет превращать его в щавелевую кислоту, которая является ядом для печени.
Алкагольдегидрогеназа превращает этиловый спирт в уксусный альдегид и степень сродства АДГ к С 2 Н 5 ОН выше, чем к этиленгликолю и на этом основан способ нейтрализации этиленгликоля.

Способ определения K m .
Построение график Михаэлиса-Ментен:
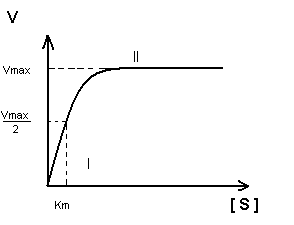
I участок – с увеличением концентрации субстрата увеличивается скорость ферментативной реакции
II участок – с увеличением концентрации субстрата скорость реакции не изменяется, т.к. все активные центры заняты.
Недостаток графика Михаэлиса-Ментен при определении K m заключается в том, что V max достигается с трудом, реакции в клетке протекают с оптимальной скоростью, а не V max .
Построение графика Лайнуэвера-Бэрка – метод обратных величин

Преимущество метода заключается в том, что прямую можно построить по двум точкам и нет необходимости определять максимальную скорость.
Зависимость скорости ферментативной реакции от концентрации фермента.
Зависимость
носит линейный характер. Скорость
химической реакции, катализируемой
данным ферментом прямопропорциональна
к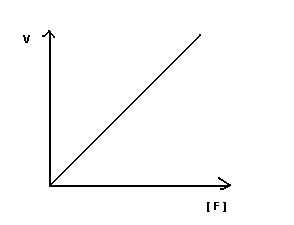 онцентрации
фермента. При этом концентрация субстрата
величина постоянная. Это обясняетя
большим количеством активных центров
при определенном количестве молекул
субстрата
онцентрации
фермента. При этом концентрация субстрата
величина постоянная. Это обясняетя
большим количеством активных центров
при определенном количестве молекул
субстрата
Билет№10
Влияние низкомолекулярных веществ на активность фенрментов.
Значение ингибирования активности ферментов.
Ингибирование лежит в основе действия лекарственных веществ и токсических агентов.
Ингибирование – один из подходов к изучению ферментативного действия (например, структуры активного центра).
ингибирование регулирует скорость и течение реакций в клетке
Ингибирование бывает 2-х видов:
Необратимое
Обратимое
Необратимое ингибирование имеет место тогда, когда присоединение ингибитора к ферменту носит необратимый характер.
Например: это действие алкилирующих агентов (подацетамид) необратимо действующих на тиогруппу ферментов. Необратимость связана с тем, что равновесие смещено вправо, в сторону образования ковалентного производного фермента:
F-S-H
+ J-CH 2 CONH 2
F-S-CH 2 -CONH 2
+ HJ
Необратимым является действие токстческих фосфоорганических соединений, которые называют нервно-паралитическими ядами, они ингибируют ацетилхолинэстеразу, участвующую в передаче нервных импульсов.
Необратимое ингибирование
Многие ингибиторы необратимо связываются с Е или ES, и, поскольку это влияет на V max , такое ингибирование относят к неконкурентному.
Ингибиторы этого типа часто ковалентно связываются с ферментом или с комплексом фермент-субстрат, необратимо изменяя нативную конфигурацию. Это объясняет токсическое действие Hg 2+ , Pb 2+ и соединений мышьяка.
На необратимом ингибировании основано действие пенициллина. Пенициллин ингибирует действие одного из ферментов, участвующих в сборке клеточной стенки бактерий. Клетки, не имеющие клеточной стенки, легко лизируются.
Действие аспирина основано на ковалентной модификации фермента. Аспирин снижает скорость синтеза простагландинов, выступая в роли ингибитора циклооксигеназного компонента эндопероксид-синтетазы. Считается, что возникновение болей, воспалений, температуры связано с простагландинами.
При интоксикации связывание яда или его вытеснение из комплекса фермент-ингибитор возможно с помощью реактиваторов, или противоядий. К ним относятся все SH – содержащие комплексоны (цистеин, димеркаптопропанол), лимонная кислота.
Обратимое ингибирование бывает 2-х видов:
Конкурентное
Неконкурентное
Обратимое конкурентное ингибирование – активность фермента восстанавливается после удаления ингибитора путем увеличения концентрации субстрата.
Отличительный признак конкурентного ингибитора – конкурентный ингибитор по структуре близок субстрату. Конкурентный ингибитор конкурирует с субстратом за активный центр фермента.
Пример: сукцинатдегидрогеназа катализирует превращение сукцината в фумарат. Конкурентным ингибитором сукцинатдегидрогеназы является малоновая кислота, которая содержит на одну группу СН 2 меньше, чем сукцинат.
СООН СООН COOH
СН 2
 СНCH 2
СНCH 2
СН 2 СН COOH
| | малоновая кислота
СООН СООН
Сукцинат и малоновая кислота являются структурными аналогами и конкурируют за активный центр фермента. (Это является подтверждением тому, что активный центр не является жестким образованием, подходящим субстрату, как «ключ-замок».)
При конкурентном ингибировании степень ингибирования фермента не зависит от абсолютной концентрации ингибитора, а от соотношения ингибитора и субстрата, если это соотношение J:S=1:50, то активность фермента ингибируется на 50%.
Действие конкурентного ингибитора снимается повышением концентрации субстрата, так как сродство фермента и субстрата выше, чем сродство фермента и ингибитора.
Кm F и S и Km F и J различны и это узнают путем построения графиков Михаэлиса-Ментен и Лайнуэвера-Бэрка
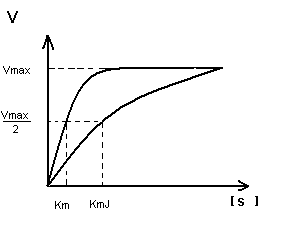

V max – одинакова
K m с ингибитором увеличивается.
Действие многих химиотерапевтических средств основано на конкурентном ингибировании. Например, сулфаниламидные препараты, используемые для лечения болезней, вызываемых микробными инфекциями. Сульфаниламидные препараты по структуре сходны с п-аминобензойной кислотой. ПАБК является предшественником в микробиологическом синтезе фолиевой кислоты, из которой кофермент, необходимый для синтеза нуклииновых кислот микроорганизмов. При введении сульфаниламидных препаратов наблюдается угнетение фермента и гибель микроорганизмов.
На конкурентном ингибировании основано применение и фторурацила, который используется при лечения рака.
Неконкурентное, обратимое ингибирование.
Действие неконкурентного ингибитора не может быть устранено увеличением концентрации субстрата.
Неконкурентный
ингибитор не
связывается
с активным центром, он может связываться
со свободным ферментом
![]() ,
либо с комплексомFS
,
либо с комплексомFS
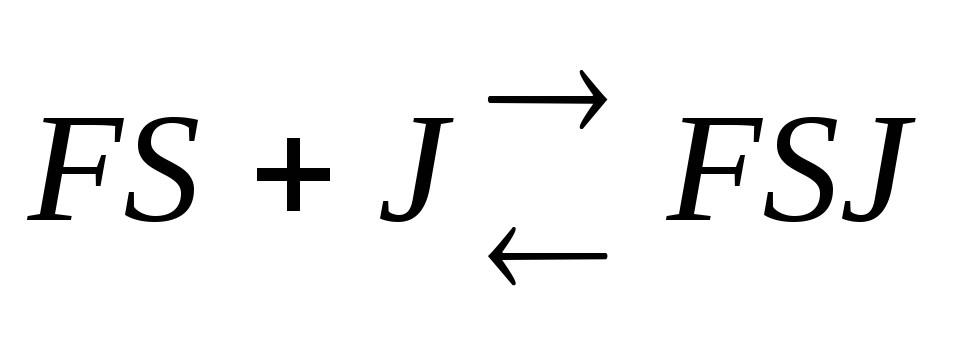 ,
либо с тем и другим, но обе формыJF
и JFS
– не активны.
,
либо с тем и другим, но обе формыJF
и JFS
– не активны.
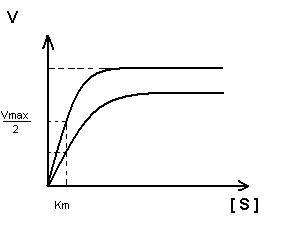

K m - не изменяется, т.к. нет связывания с активным центром.
V max – уменьшается.
Наиболее общий тип неконкурентного ингибирования имеет место при действии реагентов, обратимо связывающих SH-группы цис, входящего в каталитический центр или близко от него. Это ионы Cu 2+ , Hg 2+ , Ag + и их производные с образованием меркаптидов:
Ферменты, для активации которых необходимы ионы Ме ингибируются по такому способу агентами связывающими эти ионы:
ферро или ферроцианид.
Билет 11
Низкомолекулярные вещества, изменяющие скорость ферментативных реакций делят на 2 группы:
Активаторы – ускоряющие протекание ферментативной реакции.
Ингибиторы – замедляют протекание ферментативных реакций.
Активаторы делят на 2 группы:
В качестве активатора могут выступать коферменты или простетическая группа (в основном витамины).
Для
этой группы характерны те же закономерности,
что описаны для взаимодействия фермента
и субстрата F+S
и A+Ko
подчиняются одним закономерностям
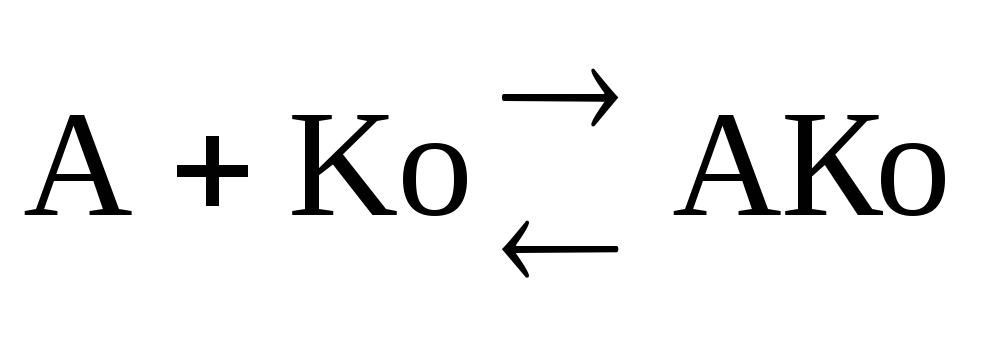
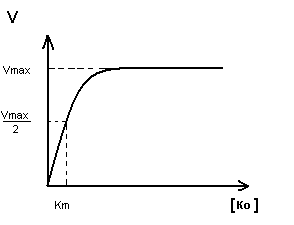
K m определяет сколько вводить Ko.
Активаторы, являющиеся связующим звеном между F и S (ориентирование фермента и субстрата) и обеспечивающим взаимодействие фермента и субстрата (F A S), взаимодействие апофермента и кофактора Апоф А Ко
Часто это ионы Ме – Со, Mn, Mg, Zn.
Активация зимогенов.
Многие пищеварительные ферменты синтезируются в неактивной форме – в виде зимогенов (пепсиноген, трипсиноген и др.).
Эти неактивные зимогены активируются после попадания в ЖКТ, благодаря этому не происходит переваривания клеток, где происходит синтез ферментов. Зимогены не активны, так как содержат дополнительные участки или группы, которые препятствуют доступу субстрата к активному центру. Удаление этой блокирующей группы протеолитическими ферментами приводит к необратимой активации зимогена.
Трипсиноген
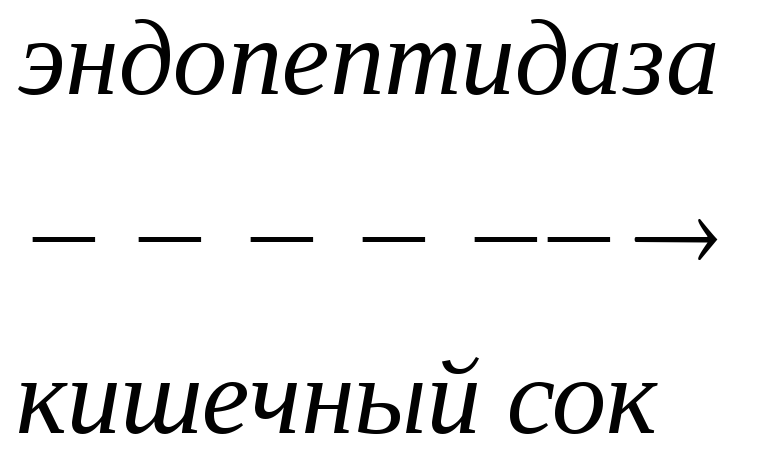

Эндопептидаза расщепляет пептидную связь между лиз в положении 6 и лей в положении 7, в результате отщепляется пептид и образуется активный трипсин.
Схема активации зимогена:
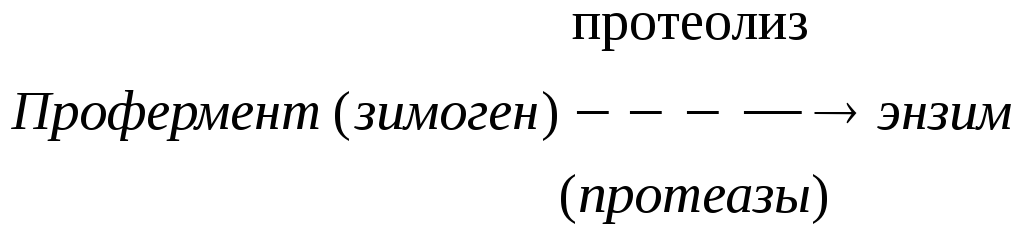
По такой схеме происходит активация целого ряда зимогенов:
Основная черта активации зимогенов заключается в каскадном (последовательном) включении зимогенов с образованием активных ферментов например 12-перстной кишки.
Регуляция путем химической модификации.
По существу это изменение химической структуры активного центра. В некоторых случаях фермент может существовать в клетке в неактивной форме или слабоактивном состоянии, но присоединение фосфатной группы при участии фосфорилирующего фермента, называемого киназой, к аминокислоте вблизи от активного центра, активирует фермент. Такая система может быть возвращена в исходное состояние под действием фермента, отщепляющего фосфатную группу.
F(неакт) Ф
Фосфотаза – Ф
Такой механизм очень важен, т.к. активность киназы и фосфатазы могут изменяться под действием гормонов.
|
Коферментная форма |
Переносимая |
Тип катализируемой |
|
|
В 1 тиамин |
ТДФ- тиаминдифосфат ТПФ (ТТФ) - тиаминпирофосфат |
Отщепление СО 2 |
Окислительное декарбоксилирование |
|
В 2 рибофла-вин |
ФМН – флавинмононуклеотид ФАД – флавинаденин-динуклеотид |
Протоны
(Н +)
и электроны ( |
Дегидрирование |
|
В 3 пантоте-новая кислота |
HSKoA – (аш эс коэнзим А) |
Ацильная группа |
Ацилирования |
|
В 5 никотин-амид |
НАД – никотинамидаде-ниндинуклеотид НАДФ – никотинамид-адениндинуклеотид фосфат |
Протоны и электроны |
Дегидрирование |
|
В 6 пиридок-саль |
ПФ – пиридоксаль фосфат |
NH 2 -группа (амино-группа) |
Трансаминирование |
|
В 7 биотин |
Коферментной формой является сам витамин |
Карбоксилирование |
|
|
В 9,10 фолиевая кислота |
ТГФК – тетрагидро-фолиевая кислота ТГФК-СН 3 – метилтетра-гидрофолиевая кислота, СН 2 ОН–ТГФК – гидроксиметилен, ТГФК-С=О – формил- |
Перенос одноуглерод-ных фрагментов: метил – СН 3 , гидрокси-метилен - СН 2 ОН, формил –С=О | |
|
В 12 кобал-амин |
СН 3 -В 12 – метил-кобаламин, ДА-В 12 – дезоксиаденозил кобаламин |
Перенос метильной группы СН 3 |
Трансметилирование |
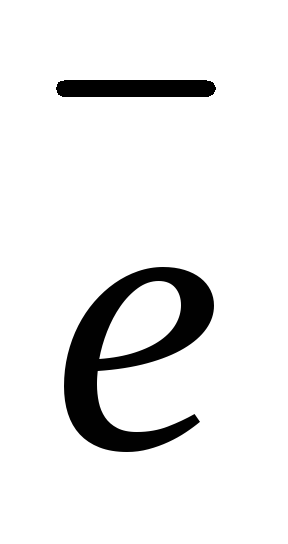 )
)Билет12
Зависимость скорости ферментативной реакции от температуры.
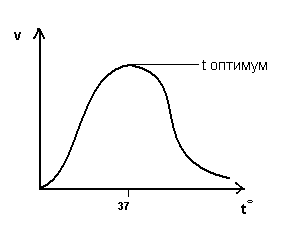 Приt=36-38 0
ферменты обладают наибольшей активностью.
Эта температура называется температурный
оптимум:
Приt=36-38 0
ферменты обладают наибольшей активностью.
Эта температура называется температурный
оптимум:
С повышением t 0 до оптимума активность ферментов повышается.
Высокие t вызывают денатурацию ферментов.
Низкие t снижают активность ферментов.
Изменение t 0 приводит к нарушению связей, закрепляющих белковую структуру ферментов (третичную, четвертичную), т.е. вызывает денатурацию.
Обратимая денатурация наблюдается при понижении t 0 . Это позволяет хранить ферменты, биологические жидкости, кровь.
Повышение температуры необратимо нарушает белковую структуру фермента. Это свойство используется при стерилизации материалов, инструментов.
Лихорадка – защитное свойство организма, т.к. происходит денатурация ферментов микроорганизмов и поэтому нецелесообразно применять жаропонижающие средства.