Медиатором в нервно мышечном синапсе является. Вещества, блокирующие передачу возбуждения. Структура и функции нервно-мышечного синапса
Синапс - это специализированная структура, которая обеспечивает передачу возбуждения с одной возбудимой структуры на другую. Термин "синапс" введен Ч.Шеррингтоном и означает "сведение", "соединение", "застежка".
Классификация синапсов. Синапсы можно классифицировать по:
их местоположению и принадлежности соответствующим структурам:
* периферические (нервно-мышечные, нейро-секреторные, рецепторнонейрональные);
* центральные (аксо-соматические, аксо-дендритные, аксо-аксональные, сомато-дендритные, сомато-соматические);
знаку их действия - возбуждающие и тормозящие;
способу передачи сигналов - химические, электрические, смешанные.
медиатору, с помощью которого осуществляется передача -холинергические, адренергические, серотонинергические, глиценергические и т.д.
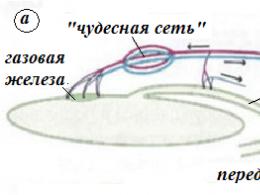
Строение синапса. Все синапсы имеют много общего, поэтому строение синапса и механизм передачи возбуждения в нем можно рассмотреть на примере нервно-мышечного синапса (рис. 7).
Синапс состоит из трех основных элементов:
* пресинаптической мембраны - (в нервно-мышечном синапсе - это утолщенная концевая пластинка);
* постсинаптической мембраны;
* синаптической щели.
Пресинаптическая мембрана - это часть мембраны мышечного окончания в области контакта его с мышечным волокном. Постсинаптическая мембрана - часть мембраны мышечного волокна. Часть постсинаптической мембраны, которая расположена напротив пресинаптической, называется субсинаптической мембраной. Особенностью субсинаптической мембраны является наличие в ней специальных рецепторов, чувствительных к определенному медиатору, и наличие хемозависимых каналов. В постсинаптической мембране, за пределами субсинаптической, имеются потенциалозависимые каналы.
Механизм передачи возбуждения в химических возбуждающих синапсах. В синапсах с химической передачей возбуждение передается с помощью медиаторов (посредников). Медиаторы - это химические вещества, которые обеспечивают передачу возбуждения в синапсах. Медиаторы в зависимости от их природы делятся на несколько групп:
* моноамины (ацетилхолин, дофамин, норадреналин, серотонин и др.);
* аминокислоты (гамма-аминомаслянная кислота - ГАМК, глутаминовая кислота, глицин и др.);
* нейропептиды (вещество Р, эндорфины, нейротензин, АКТГ, ангиотензин, вазопрессин, соматостатин и др.).
Медиатор в молекулярном виде находится в пузырьках пресинаптического утолщения (синаптической бляшке), куда он поступает:
* из околоядерной области нейрона с помощью быстрого аксонального транспорта (аксотока);
* за счет синтеза медиатора, протекающего в синаптических терминалях из продуктов его расщепления;
* за счет обратного захвата медиатора из синаптической щели в неизменном виде.
Когда по аксону к его терминалям приходит возбуждение, пресинаптическая мембрана деполяризуется, что сопровождается поступлением ионов кальция из внеклеточной жидкости внутрь нервного окончания. Поступившие ионы кальция активируют перемещение синаптических пузырьков к пресинаптичской мембране, их соприкосновение и разрушение (лизис) их мембран с выходом медиатора в синаптическую щель. В ней медиатор диффундирует к субсинаптической мембране, на которой находятся его рецепторы. Взаимодействие медиатора с рецепторами приводит к открытию преимущественно каналов для ионов натрия. Это приводит к деполяризации субсинаптической мембраны и возникновению так называемого возбуждающего постсинаптического потенциала (ВПСП). В нервно-мышечном синапсе ВПСП называется потенциалом концевой пластинки (ПКП). Между деполяризованной субсинаптической мембраной и соседними с ней участками постсинаптическои мембраны возникают местные токи, которые деполяризуют мембрану. Когда они деполяризуют мембрану до критического уровня, в постсинаптической мембране мышечного волокна возникает потенциал действия, который распространяется по мембранам мышечного волокна и вызывает его сокращение.
Химические тормозные синапсы. Эти синапсы по механизму передачи возбуждения сходны с синапсами возбуждающего действия. В тормозных синапсах медиатор (например, глицин) взаимодействует с рецепторами субсинаптической мембраны и открывает в ней хлорные каналы, это приводит к движению ионов хлора по концентрационному градиенту внутрь клетки и развитию гиперполяризации на субсинаптической мембране. Возникает так называемый тормозной постсинаптический Потенциал (ТПСП).
Ранее полагали, что каждому медиатору соответствует специфическая реакция постсинаптической клетки - возбуждение или торможение в той или иной форме. В настоящее время установлено, что одному медиатору чаше всего соответствует.не один, а несколько, различных рецепторов. Например, ацетилхолин в нервно-мышечных синапсах скелетных мышц действует на Н-холинорецепторы (чувствительные к никотину), которые открывают широкие каналы для натрия (и калия), что порождает ВПСП (ПКП). В ваго-сердечных синапсах тот же ацетилхолин действует на М-холинорецепторы (чувствительные к мускарину), открывающие селективные каналы для ионов калия, поэтому здесь генерируется тормозной постсинаптический потенциал (ТПСП). Следовательно, возбуждающий или тормозной характер действия медиатора определяется свойствами субсинаптической мембраны (точнее, видом рецептора), а не самого медиатора.
Физиологические свойства химических синапсов.
Синапсы с химической передачей возбуждения обладают рядом общих свойств:
* Возбуждение через синапсы проводится только в одном направлении (односторонне). Это обусловлено строением синапса: медиатор выделяется только из пресинаптического утолщения и взаимодействует с рецепторами субсинаптической мембраны;
* передача возбуждения через синапсы осуществляется медленнее, чем по -нервному волокну - синаптическая задержка;
* передача возбуждения осуществляется с помощью специальных химических посредников - медиаторов;
* в синапсах происходит трансформация ритма возбуждения;
* синапсы обладают низкой лабильностью;
* синапсы обладают высокой утомляемостью;
* синапсы обладают высокой чувствительностью к химическим (в том числе и фармакологическим) веществам.
Электрические синапсы возбуждающего действия. Кроме синапсов с химической передачей возбуждения преимущественно в центральной нервной системе (ЦНС) встречаются синапсы с электрической передачей. Возбуждающим электрическим синапсам свойственны очень узкая синаптическая щель и очень низкое удельное сопротивление сближенных пре- и постсинаптических мембран, что обеспечивает эффективное прохождение локальных электрических токов. Низкое сопротивление, как правило, связано с наличием поперечных каналов, пересекающих обе мембраны, т. е. идущих из клетки в клетку (щелевой контакт). Каналы образуются белковыми молекулами (полумолекулами) каждой из контактирующих мембран, которые соединяются комплементарно. Эта структура легко проходима для электрического тока.
Схема передачи возбуждения в электрическом синапсе: ток, вызванный пресинаптическим потенциалом действия, раздражает постсинаптическую мембрану, где возникает ВПСП и потенциал действия.
Поперечные каналы объединяют клетки не только электрически, но и химически, так как они проходимы для многих низкомолекулярных соединений. Поэтому возбуждающие электрические синапсы с поперечными каналами формируются, как правило, между клетками одного типа (например, между клетками сердечной мышцы).
Общими свойствами возбуждающих электрических синапсов являются:
* быстродействие, (значительно превосходит таковое в химических синапсах);
* слабость следовых эффектов при передаче возбуждения (в результате этого в них практически невозможна суммация последовательных сигналов);
* высокая надежность - передачи возбуждения.
Возбуждающие электрические синапсы могут возникать при благоприятных условиях и исчезать при неблагоприятных. Например, при повреждении одной из контактирующих клеток ее электрические синапсы с другими клетками ликвидируются. Это свойство называется пластичностью.
Электрические синапсы могут быть с односторонней и двусторонней передачей возбуждения.
Электрический тормозной синапс . Наряду с электрическими синапсами возбуждающего действия могут встречаться электрические тормозные синапсы. Примером такого синапса может служить синапс, который образует нервное окончание на выходном сегменте маутнеровского нейрона у рыб. Тормозящее влияние возникает за счет действия тока, вызванного потенциалом действия пресинаптической мембраны. Пресинаптический потенциал вызывает значительную гиперполяризацию сегмента и гиперполяризующий ток мгновенно тормозит генерацию потенциала действия в начальном сегменте аксона.
В смешанных синапсах пресинаптический потенциал действия создает ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны неплотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ .
ФУНКЦИИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ . Организм человека представляет собой сложную высокоорганизованную систему, состоящую из функционально связанных между собой клеток, тканей, органов и их систем.
Эту взаимосвязь (интеграцию) функций, их согласованное функционирование обеспечивает центральная нервная система (ЦНС). ЦНС регулирует все процессы, протекающие в организме, поэтому с ее помощью происходят наиболее адекватные изменения работы различных органов, направленные на обеспечение той или иной его деятельности.
ЦНС также осуществляет связь организма с внешней средой, путем анализа и синтеза поступающей к ней разнообразной информации от рецепторов. Она выполняет функции регулятора поведения, необходимого в конкретных условиях существования. Это обеспечивает адекватное приспособление к окружающему миру. Кроме того с функциями ЦНС связаны процессы, лежащие в основе психической деятельности человека.
МЕТОДЫ ИССЛЕДОВАНИЯ ФУНКЦИЙ ЦНС . Интенсивное развитие функций ЦНС обусловило переход от описательных методов изучения функций различных отделов мозга к экспериментальным методам. Многие методы, используемые для изучения функции ЦНС, применяются в сочетании друг с другом.
Метод разрушения (экстерпации) различных отделов ЦНС. С помощью этого метода можно установить какие функции ЦНС выпадают после оперативного вмешательства и какие сохраняются. Данный методический прием давно испольуется в экспериментально-физиологических исследованиях.
Метод перерезки дает возможность изучить значение в деятельности того или иного отдела ЦНС влияний, поступающих от других ее отделов. Перерезка производится на различных уровнях ЦНС. Полная перерезка, например, спинного мозга или ствола мозга разобщает вышележащие отделы ЦНС от нижележащих и позволяет изучить рефлекторные реакции, которые осуществляются нервными центрами, расположенными ниже места перерезки. Перерезка и локальное повреждение отдельных нервных центров производится не только в условиях эксперимента, но и в нейрохирургической клинике в качестве лечебных мероприятий.
Метод раздражения позволяет изучить функциональное значение различных образований ЦНС. При раздражении (химическом, электрическом, механическом и т.д.) определенных структур мозга можно наблюдать возникновение, особенности проявления и характер распространения процессов возбуждения.
Электроэнцефалография - метод регистрации суммарной электрической активности различных отделов головного мозга. Впервые запись электрической активности мозга была осуществлена В. В. Правдич-Неминским с помощью электродов, погруженных в мозг. Бергер зарегистрировал потенциалы мозга с поверхности черепа и назвал запись колебаний потенциалов мозга электроэнцефалограммой (ЭЭГ-ма).
Частота и амплитуда колебаний может меняться, но в каждый момент времени в ЭЭГ-ме преобладают определенные ритмы, которые Бергер назвал альфа-, бета-, тета- и дельта-ритмами. Альфа-ритм характеризуется частотой колебаний 8-13 Гц, амплитуда 50 мкВ. Этот ритм лучше всего выражен в затылочной и теменной области коры и регистрируется в условиях физического и умственного покоя при закрытых глазах. Если глаза открыть, то альфа-ритм сменяется более быстрым бета-ритмом. Бета-ритм характеризуется частотой колебаний 14-50 Гц и амплитудой до 25 мкВ. У некоторых людей альфа-ритм отсутствует, и поэтому в покое регистрируется бета-ритм. В связи с этим различают бета-ритм 1 с частотой колебаний 16-20 Гц, он характерен для состояния покоя и регистрируется в лобной и теменной областях. Бета-ритм 2 с частотой 20-50 Гц и характерен он для состояния интенсивной деятельности мозга. Тета-ритм представляет собой колебания с частотой 4-8 Гц и амплитудой 100-150 мкВ. Этот ритм регистрируется в височной и теменной областях при психомоторной активности, при стрессе, во время сна, при гипоксии и легком наркозе. Дельта-ритм характеризуется медленными колебаниями потенциалов с частотой 0,5-3,5 Гц, амплитудой 250-300 мкВ. Этот ритм регистрируется во время глубокого сна, при глубоком наркозе, при гипоксии.
ЭЭГ метод используется в клинике с диагностической целью. Особенно широкое применение этот метод нашел в нейрохирургической клинике для определения локализации опухолей мозга. В неврологической клинике этот метод находит применение при определении локализации эпилептического очага, в психиатрической клинике - для диагностики расстройств психики. В хирургической клинике ЭЭГ используется для тестирования глубины наркоза.
Метод вызванных потенциалов - регистрация электрической активности определенных структур мозга при стимуляции рецепторов, нервов, подкорковых структур. Вызванные потенциалы (ВП) чаще всего представляют собой трехфазные колебания ЭЭГ-мы, сменяющие друг друга: позитивное, негативное, второе (позднее) позитивное колебание. Однако, они могут иметь и более сложную форму. Различают первичные (ПО) и поздние или вторичные (ВО) вызванные потенциалы. ВП - это фрагмент ЭЭГ-мы, записанный в момент стимуляции мозга и имеет ту же природу, что и электроэнцефалограмма.
Метод ВП находит применение в неврологии и в нейрофизиологии. С помощью ВП можно проследить онтогенетическое развитие проводящих путей мозга, провести анализ локализации представительства сенсорных функций, провести анализ связей между структурами мозга, показать количество переключений на пути распространения возбуждения и т.д.
Микроэлектродный метод применяется для изучения физиологии отдельного нейрона, его биоэлектрической активности как в состоянии покоя, так и при различных воздействиях. Для этих целей используются специально изготовленные стеклянные или металлические микроэлектроды, диаметр кончика которых составляет 0,5-1,0 мкм или чуть больше. Стеклянные микроэлектроды представляюют собой микропипетки, заполненные раствором электролита. В зависимости от расположения микроэлектрода различают два способа отведения биоэлектрической активности клеток - внутриклеточное и внеклеточное.
Внутриклеточное отведение позволяет регистрировать и измерять:
* мембранный потенциал покоя;
* постсинаптические потенциалы (ВПСП и ТПСП);
* динамику перехода местного возбуждения в распространяющееся;
* потенциал действия и его компоненты.
Внеклеточное отведение дает возможность регистрировать:
* спайковую активность как отдельных нейронов, так и, в основном, их групп, расположенных вокруг электрода.
Для точного определения положения различных структур головного мозга и для введения в них различных микропредметов (электроды, термопары, пипетки и др.) широкое применение как в электрофизиологических исследованиях, так и в нейрохирургической клинике нашел стереотаксический метод. Его использование основано на результатах детальных анатомических исследований расположения различных структур головного мозга относительно костных ориентиров черепа. По данным таких исследований созданы специальные стереотаксические атласы как для различных видов животных, так и для человека. В настоящее время стереотаксический метод находит широкое применение в нейрохирургической клинике для следующих целей:
* разрушения структур мозга с целью ликвидации состояний гиперкинеза, неукротимой боли, некоторых психических расстройств, эпилептических нарушений и др. ;
* выявления патологических эпилептогенных очагов;
* введения радиоактивных веществ в опухоли мозга и для разрушения этих опухолей;
* коагуляции аневризм мозговых сосудов;
* осуществления лечебных электростимуляций или торможений структур мозга.
СТРОЕНИЕ ЦНС . Структурно-функциональной единицей ЦНС является нейрон (нервная клетка). Он состоит из тела (сомы) и отростков - многочисленных дендритов и одного аксона. Дендриты обычно сильно ветвятся и образуют множество синапсов, с другими клетками, что определяет их ведущую роль в восприятии нейроном информации. Аксон, начинается от тела клетки аксонным холмиком, функцией которого является генерация нервного импульса, который по аксону проводится к другим клеткам. Аксон сильно ветвится, образуя множество коллатералей, терминали которых образуют синапсы с другими клетками. Мембрана аксона в области синапса содержит специфические рецепторы, способные реагировать на различные медиаторы или нейромодуляторы. Поэтому процесс выделения медиатора пресинаптическими окончаниями может эффективно регулироваться другими нейронами. Кроме того, мембрана окончаний содержит большое число потенциалозависимых кальциевых каналов, через которые ионы кальция поступают внутрь окончания при его возбуждении.
В большинстве центральных нейронов ПД возникает в области мембраны аксонного холмика, возбудимость которой в два раза выше других участков и отсюда возбуждение распространяется по аксону и телу клетки. Такой способ возбуждения нейрона важен для осуществления его интегративной функции, т. е. способности суммировать влияния, поступающие на нейрон по разным синаптическим путям. Степень возбудимости разных участков нейрона неодинакова, она самая высокая в области аксонного холмика, в области тела нейрона она значительно ниже, и самая низкая у дендритов.
Помимо нейронов в ЦНС имеются глиальные клетки, занимающие половину объема мозга. Периферические аксоны также окружены оболочкой из глиальных клеток -ивановских клеток. Нейроны и глиальные клетки разделены межклеточными щелями, которые сообщаются друг с другом и образуют заполненное жидкостью межклеточное пространство нейронов и глии. Через это пространство происходит обмен веществами между нервными и глиальными клетками. Функции клеток глии многообразны:
* они являются для нейронов опорным, защитным и трофическим аппаратом, поддерживают определенную концентрацию ионов калия и кальция в межклеточном пространстве;
* активно поглощают нейромедиаторы, ограничивая таким образом время их
действия и другие функции.
АКСОННЫЙ ТРАНСПОРТ . Аксоны, помимо функции проведения возбуждения, являются каналами для транспорта различных веществ. Белки и медиатор, синтезированные в теле клетки, органеллы и другие вещества могут перемещаться по аксону к его окончанию. Это перемещение веществ получило название аксонного транспорта. Существует два его вида - быстрый и медленный аксонный транспорт.
БЫСТРЫЙ АКСОННЫЙ ТРАНСПОРТ - это транспорт везикул, митохондрий и некоторых белковых частиц от тела клетки к окончаниям аксона (антероградный транспорт) со скоростью 250-400 мм/сут. Он осуществляется специальным транспортным механизмом - при помощи микротрубочек и нейрофиламентов и сходен с механизмом мышечного сокращения.
Быстрый аксонный транспорт от терминалей аксона к телу клетки, или ретроградный, перемещает лизосомы, везикулы, возникающие в окончаниях аксона в ходе пиноцитоза, например, ацетилхолинэстеразы, некоторых вирусов, токсинов и др. со скоростью 220 мм/сут. Скорость быстрого антероградного и ретроградного транспорта не зависит от типа и диаметра аксона.
МЕДЛЕННЫЙ АКСОННЫЙ ТРАНСПОРТ обеспечивает перемещение со скоростью 1-4 мм/сут. белков и структур цитоплазмы (микротрубочек, нейрофиламентов, РНК, транспортных и канальных мембранных белков и т. д.) в дистальном направлении за счет интенсивности синтетических процессов в перикарионе. Медленный аксонный транспорт имеет особое значение в процессах роста и регенерации отростков нейрона.
РАЗВИТИЕ РЕФЛЕКТОРНОЙ ТЕОРИИ.
Основным механизмом деятельности ЦНС является рефлекс. Рефлекс - это ответная реакция организма на действия раздражителя, осуществляемая с участием ЦНС и направленная на достижение полезного результата.
Рефлекс в переводе с латинского языка означает "отражение"-. Впервые термин "отражение" или "рефлектирование" был применен Р. Декартом (1595-1650) для характеристики реакций организма в ответ на раздражение органов чувств. Он первым высказал мысль о том, что все проявления эффекторной активности организма вызываются вполне реальными физическими факторами. После Р. Декарта представление о рефлексе было развито чешским исследователем Г. Прохазкой, который развил учение об отражательных действиях. В это время уже было отмечено, что у спинальных животных движения наступают в ответ на раздражение определенных участков кожи, а разрушение спинного мозга ведет к их исчезновению.
Дальнейшее развитие рефлекторной теории связано с именем И. М. Сеченова. В книге "Рефлексы головного мозга" он утверждал, что все акты бессознательной и сознательной жизни по природе происхождения являются рефлексами. Это была гениальная попытка ввести физиологический анализ в психические процессы. Но в то время не существовало методов объективной оценки деятельности мозга, которые могли бы подтвердить это предложение И. М. Сеченова. Такой объективный метод был разработан И. П. Павловым - метод условных рефлексов, с помощью которого он доказал, что высшая нервная деятельность организма, так же как и низшая, является рефлекторной.
Структурной основой рефлекса, его материальным субстратом (морфологической основой) является рефлекторная дуга - совокупность морфологических структур, которая обеспечивает осуществление рефлекса (путь, по которому проходит возбуждение при осуществлении рефлекса).
В основе современное представления о рефлекторной деятельности лежит понятие полезного приспособительного результата, ради которого совершается любой рефлекс. Информация о достижении полезного приспособительного результата поступает в ЦНС по звену обратной связи в виде обратной афферентации, которая является обязательным компонентом рефлекторной деятельности. Принцип обратной афферентации введен в рефлекторную теорию П. К. Анохиным. Таким образом, по современным представлениям структурной основой рефлекса является не рефлекторная дуга, а рефлекторное кольцо, состоящее из следующих компонентов (звеньев):
* рецептор;
* афферентный нервный путь;
* нервный центр;
* эфферентный нервный путь;
* рабочий орган (эффектор);
* обратная афферентация (рис. 8).
Анализ структурной основы рефлекса проводится путем последовательного выключения отдельных звеньев рефлекторного кольца (рецептора, афферентного и эфферентного пути, нервного центра). При выключении любого звена рефлекторного кольца рефлекс исчезает. Следовательно, для осуществления рефлекса необходима целостность всех звеньев его морфологической основы.
Клетки ЦНС имеют многочисленные связи Друг с другом, поэтому нервная система человека может быть представлена как система нейронных цепей (нейронных сетей), передающих возбуждение и формирующих торможение. В этой нервной сети возбуждение может распространяться от одного нейрона на многие другие нейроны. Процесс распространения возбуждения от одного нейрона на многие другие нейроны получил название иррадиации возбуждения или дивергентного принципа распространения возбуждения.
Различают два вида иррадиации возбуждения :
* направленная или системная иррадиация когда возбуждение распространяется по определенной системе нейронов и формирует координированную приспособительную деятельность организма;
* бессистемная или диффузная (ненаправленная) иррадиация, хаотичное распространение возбуждения, при котором координированная деятельность невозможна (рис. 9).
В ЦНС к одному нейрону могут сходиться возбуждения от различных источников. Эта способность возбуждений сходиться к одним и тем же промежуточным и конечным нейронам получила название конвергенции возбуждений (рис. 9).
Пресинаптическая мембрана нервно-мышечного синапса представляет собой часть мембраны пресинаптического окончания аксона мотонейрона, ограничивающая синаптическую щель. Через нее осуществляется выделение (экзоитоз) медиатора в синаптическую щель. Медиатор пресинаптического окончания содержится в синаптических пузырьках(везикулах) диаметром 40 нм. Они образуются в комплесе Гольджи, с помощью быстрого прямого аксонного транспорта доставляются в пресинаптическое окончание и там заполняются медиатором и АТФ. В нервно-мышечном синапсе медиатором является ацетилхолин, который образуется из ацетилкоэнзима А и холина под действием фермента холинацетилтрансферазы. Везикулы расположены преимущественно вблизи периодических утолщений пресинаптической мембраны, называемых активными зонами. В неактивном синапсе везикулы с помощью белка синапсина связаны с белками цитоскелета,что обеспечивает им иммобилизацию и резервирование. Важными структурами пресинаптического окончания являются митохондрии, осуществляющие энергетическое обеспечение процесса синаптической передачи, цистерны гладкой ЭПС, содержащие депонированный ион Са, микротрубочки и микрофиламенты, участвующие во внутриклеточном передвижении везикул.
Синаптическая щель в нервно-мышечном синапсе имеет ширину в среднем 50 нм. Она содержит межклеточную жидкость и мукополисахаридное плотное вещество в виде полосок, мостиков, которое называется базальной мембраной и содержит ацетилхолинэстеразу.
Постсинаптическая мембрана содержит рецепторы,способные связывать молекулы медиатора. Ее особенностью является наличие мелких складок, которые образуют карманы, открывающиеся в синаптическую щель.
Таким образом, основными этапами передачи возбуждения в нервно-мышечном синапсе являются:
1) возбуждение мотонейрона, распространение потенциала действия на пресинаптическую мембрану;
2) повышение проницаемости пресинаптической мембраны для ионов кальция, ток кальция в клетку, повышение концентрации кальция в пресинаптическомокончаниии;
3) слияние синаптических пузырьков с пресинаптической мембраной в активной зоне, экзоцитоз, поступление медиатора в синаптическую щель;
4) диффузия ацетилхолина к постсинаптической мембране, присоединение его к N-холинорецепторам, открытие хемозависимых ионных каналов;
5) преобладающий ионный ток натрия через хемозависимые каналы, образование надпорогового потенциала концевой пластинки;
6) возникновение потенциалов действия на мышечной мембране;
7) ферментативное расщепление ацетилхолина, возвращение продуктов расщепления в окончание нейрона, синтез новых порций медиатора.
Миорелаксанты- лекарственные средства, снижающие тонус скелетной мускулатуры с уменьшением двигательной активности вплоть до полного обездвиживания.
Механизм действия - блокада Н-холинорецепторов в синапсах прекращает подачу нервного импульса к скелетным мышцам, и мышцы перестают сокращаться. Расслабление идет снизу вверх, от кончиков пальцев ног до мимических мышц. Последней расслабляется диафрагма. Восстановление проводимости идет в обратном порядке.
По особенностям взаимодействия с рецепторами миорелаксанты делятся на две группы:
Деполяризирующие миорелаксанты - при контакте с рецепторами вызывают стойкую деполяризацию мембраны синапса, сопровождающуюся кратковременным хаотичным сокращением мышечных волокон (миофасцикуляции), переходящим в миорелаксацию. При стойкой деполяризации нервно-мышечная передача прекращается. Миорелаксациянепродолжительна, происходит за счет удержания открытыми мембранных каналов и невозможности реполяризации. Метаболизируютсяпсевдохолинэстеразой, выводятся почками
Недеполяризирующиемиорелаксанты - блокируют рецепторы и мембранные каналы без их открытия, не вызывая деполяризацию
20в. Физиологические свойства и особенности гладких мышц
Основные функции мышечной ткани:
1.двигательная – обеспечение движения
2.статическая – обеспечение фиксации, в том числе и в определенной позе
3.рецепторная – в мышцах имеются рецепторы, позволяющие воспринимать собственные движения
4.депонирующая – в мышцах запасаются вода и некоторые питательные вещества.
Физиологические свойства:
Сократимость. Сокращение гладкой мышцы определяется особым характером распостранения возбуждения. Группа клеток взаимодействующих посредством нексусов и своих электрических полей, формирует пучок, который является структурно-функциональной единицей гладкой мышцы и сокращается как единое целое.большая продолжительность сокращения; Время сокращения гладкомышечных волокна в несколько сотен раз больше, чем поперечно. Благодаря этому гладкие мышцы приспособлены к длительному сокращению без больших затрат энергии и медленно устают;
Спонтанная миогенная активность. В отличие от скелетных мышц гладкие мышцы желудка, кишечника, матки, мочеточников, кровеносных сосудов и других внутренних органов развивают спонтанно-тетаноподобные сокращения.Эта спонтанная активность возникает в особых мышечных клетках, которые выполняют функцию водителя ритма, то есть обладают способностью до автоматизма. От этих клеток ПД распространяется со скоростью примерно 0.1 м/с через Нексус на соседние волокна и охватывает всю мышцу. Например, перистальтические сокращения желудка возникают с частотой 3 раза за 1 мин, сегментарные и маятниковые движения в толстом кишечнике - с частотой -20 раз за 1 мин.
Пластичность - способность сохранять длину, приобретенную при растяжении, без изменения напряжения. Это свойство имеет очень большое значение для нормальной деятельности внутренних органов, например мочевого пузыря.
высокая чувствительность к физиологически активных веществ, в частности к медиаторам вегетативной нервной системы - ацетилхолина, а также - серотонина, брадикинина, простагландинов. Указанные биологически активные вещества могут как возбуждать, так и тормозить гладкомышечные волокна. Это зависит от того, какой процесс - деполяризацию, или гиперполяризацию вызывает данное вещество на мембране клетки. Так, например, ацетилхолин вызывает сокращение гладких мышц большинства органов, но способствует расслаблению стенок сосудов некоторых органов. Характер ответы гладких мышц на действие физиологически активного вещества зависит от того, ионные каналы она открывает в свою очередь детерминировано спецификой мембранных рецепторов.
Возбудимость. Потенциал покоя 60-70 мВ. Для миоцитов,обладающих спонтанной элекрической активностью 30-60. Положительный пик ПД меньше, чем в поперечнополосатых меньше, чем в поперечнополосатых мышечных волокнах и достигает 10-15 мВ. Длительность ПД колеблется от 25 мс до 1 с. В процессе формирования потенциала покоя играют роль не только ионы К +, но и Са) Взаимодействие актина и миозина в гладкомышечных волокнах также активируется ионами Са 2, но они попадают в клетки не с сарко-ретикулума, а транспортируются туда с межклеточного среды. Деполяризация мембраны обусловлена открытием кальциевых каналов и диффузией ионов кальция в клетку.
Проводимость. Проведение возбуждения по гладкомышечному миоциту непрерывное. Однако изолированно отдельные гладкомышечные клетки не возбуждаются и не сокращаются. Взаимодействие между отдельными миоцитами осуществляется благодаря щелевидным контактам, обладающим низким электрическим сопротивлением. Благодаря этому электрическое поле 1 клетки обеспечивает возбуждение другой. Скорость распространения ПД в пределах пучка составляет 5-10 м/с.
Автоматия присуща клеткам-водителя ритма (пейсмекерам). В ее основе лежит спонтанно возникающая медленная деполяризация – при достижении критического потенциала возникает ПД. Эта деполяризация преимущественно обусловлена диффузией ионов кальция в клетку.
2.Рефлекторная дуга – совокупность структур при помощи которой осуществляется рефлекс. Схематично рефлекторную дугу можно изобразить из 5 звеньев.
1. Воспринимающее звено (рецептор) обеспечивает восприятие изменений внешней и внутренней среды организма посредством трансформации энергии раздражения в рецепторный потенциал.
Рефлексогенная зона – совокупность рецепторов, раздражение которых вызывает рефлекс. При любом раздражении возникают рецепторные потенциалы, обеспечивающие посылку н.и. в ЦНС с помощью 2 звена.
2. Афферентное звено
Роль: передача сигнала в цнс к третьему звену рефлекторной дуги. Для соматической нервной системы - это афферентный нейрон с его отростками, тело его расположено в спинномозговых ганглиях или ганглиях черепных нервов. Импульс по дендриту афферентного нейрона, затем по его аксону, далее в цнс.
3. Управляющее звено – совокупность центральных (для ВНС и ПНС) нейронов, формирующих ответную реакцию организма.
4. Эфферентное звено – аксон эффекторного нейрона (для соматической н.с. мотонейрона).
5. Эффектор (рабочий орган) эффекторным нейроном соматической н.с. является мотонейрон
Классификация рефлексов
1. По условиям появления рефлексов в онтогенезе
a) Врожденные (безусловные)
b) Приобретенные
Врожденные могут быть соматическими (с помощью сомат. н.с., в качестве эффектора – скелетная мускулатура) и вегетативными (с помощью вегет. н.с.)
2. По биологическому значению
a) Гомеастатические (регуляция функций внутр. органов; работа сердца; секреция и моторика ЖКТ – пищевые рефлексы.)
b) Защитные (оборонительные)
c) Половые
d) ориентировочный рефлекс.
3. В зависимости от числа синапсов
В центральной части рефлекторной дуги различают.
а) моносинаптические (рефлекс на растяжение четырехглавной мышцы - коленный разгибательный рефлекс, при ударе по сухожилию
б) полисинаптические (участвует несколько последовательно включенных нейронов ЦНС
4. по рецепторам, раздражение которых вызывает ответную реакцию.
а) экстероцептивные
б) интероцептивные
в) проприоцептивные (используются в клинической практике для оценки состояния возбудимости ЦНС и для диагностики.
5. по локализации рефлекторной дуги
А) центральные (дуга через ЦНС)
б) периферические ревлексы (дуга замыкается вне ЦНС)
в) по отношению к физиологическим системам
|
ПРИЗНАКИ |
ВЕГЕТАТИВНАЯ |
СОМАТИЧЕСКАЯ |
|
Органы-мишени |
Гладкие мышцы, миокард, железы, жировая ткань, органы иммунитета |
Скелетные мышцы |
|
Паравертебральные, Превертебральные и Органные |
Локализованы в ЦНС |
|
|
Число эфферентных нейронов | ||
|
Эффект стимуляции |
Возбуждающий или Подавляющий |
Возбуждающий |
|
Типы нервных волокон |
Тонкие миелиновые или немиелиновые, медленные |
миелиновые быстрые |
Синапс (греч. synapsis - соединение) - это специализированная структура, обеспечивающая передачу сигнала от клетки к клетке. Посредством синапса реализуется действие многих фармакологических препаратов.
Структурно-функциональная организация. Каждый синапс имеет пре - и постсинаптическую мембраны и синаптическую щель (рис. 17).
Рис. 17. Нервно-мышечный синапс скелетной мышцы: 1 – ветвь аксона; 2 – пресинаптическое окончание аксона; 3 – митохондрия; 4 – синаптические пузырьки, содержащие ацетилхолин; 5 – синаптическая щель; 6 – молекулы медиатора в синаптической щели; 7 – постсинаптическая мембрана мышечного волокна с N-холинорецепторами
Пресинаптическая мембрана нервно-мышечного синапса представляет собой часть мембраны пресинаптического окончания аксона мотонейрона. Через нее осуществляется выброс (экзоцитоз) медиатора (лат. mediator - посредник) в синаптическую щель. В нервно-мышечном синапсе медиатором является ацетилхолин. Медиатор пресинаптического окончания содержится в синаптических пузырьках (везикулах), диаметр которых составляет около 40 нм. Они образуются в комплексе Гольджи, с помощью быстрого аксонного транспорта доставляются в пресинаптическое окончание, где заполняются медиатором и АТФ. В пресинаптическом окончании содержится несколько тысяч везикул, в каждой из которых имеется от 1 тыс. до 10 тыс. молекул химического вещества.
Постсинаптическая мембрана (концевая пластинка в нервно-мышечном синапсе) - это часть клеточной мембраны иннервируемой мышечной клетки, содержащая рецепторы, способные связывать молекулы ацетилхолина. Особенность этой мембраны: множества мелких складок, увеличивающих ее площадь и количество рецепторов на ней до 10-20 млн в одном синапсе.
Синаптическая щель в нервно-мышечном синапсе имеет ширину в среднем 50 нм. Она содержит межклеточную жидкость, ацетилхолинэстеразу и мукополисахаридное плотное вещество в виде полосок, мостиков, в совокупности образующих базальную мембрану, соединяющую пре- и постсинаптическую мембраны.
Механизмы синаптической передачи включают три основных этапа (рис. 18).
Рис. 18. Механизм проведения импульса через химический синапс: 1-8 – этапы процесса (Чеснокова, 2007)
Первый этап - процесс выброса медиатора в синаптическую щель, который запускается посредством ПД пресинаптического окончания. Деполяризация его мембраны ведет к открытию потенциалуправляемых Са-каналов. Са 2+ входит в нервное окончание согласно электрохимическому градиенту. Часть медиатора в пресинаптическом окончании локализуется на пресинаптической мембране изнутри. Са 2+ активирует экзоцитозный аппарат пресинапса, представляющий собой совокупность белков (синапсин, спектрин и др.), пресинаптического окончания, активация которых обеспечивает выброс ацетилхолина посредством экзоцитоза в синаптическую щель. Количество высвобождаемого ацетилхолина из пресинаптического окончания пропорционально в четвертой степени количеству поступившего туда Са 2+ . На один ПД из пресинаптического окончания нервно-мышечного синапса выбрасывается 200-300 квантов (везикул) медиатора.
Второй этап - диффузия ацетилхолина в течение 0,1-0,2 мс к постсинаптической мембране и действие его на N-холинорецепторы (стимулируются также никотином, вследствие чего и получили свое название). Удаление ацетилхолина из синаптической щели осуществляется путем разрушения его под действием ацетилхолинэстеразы, расположенной в базальной мембране синаптической щели, в течение нескольких десятых долей миллисекунды. Около 60% холина захватывается обратно пресинаптическим окончанием, что делает синтез медиатора более экономичным, часть ацетилхолина рассеивается. В промежутках между ПД из пресинаптического окончания происходит спонтанное выделение 1- 2 квантов медиатора в синаптическую щель в течение 1 с, формируя так называемые миниатюрные потенциалы (0,4-0,8 мВ). Они поддерживают высокую возбудимость иннервируемой клетки в условиях функционального покоя и выполняют трофическую роль, а в ЦНС - способствуют поддержанию тонуса ее центров.
Третий этап - взаимодействие ацетилхолина с N-холинорецепторами постсинаптической мембраны, в результате чего открываются ионные каналы на 1 мс и, вследствие преобладания входа N + в клетку, происходит деполяризация постсинаптичедкой мембраны (концевой пластинки). Эту деполяризацию в нервно-мышечном синапсе называют потенциалом концевой пластинки (ПКП) (рис. 19).
Особенностью нервно-мышечного синапса скелетного мышечного волокна является то, что при одиночной его активации формируется ПКП большой амплитуды (30-40 мВ), электрическое поле которого вызывает генерацию ПД на мембране мышечного волокна вблизи синапса. Большая амплитуда ПКП обусловлена тем, что нервные окончания делятся на многочисленные веточки, каждая из которых выбрасывает медиатор.

Рис. 19. Потенциал концевой пластинки (Шмидт, 1985): КП – критический потенциал; ПД – потенциал действия; А – ПКП в нормальной мышце; Б – ослабленный ПКП в курарезированной мышце; стрелками указан момент нанесения стимула
Характеристика проведения возбуждения в химических синапсах . Одностороннее проведение возбуждения от нервного волокна к нервной или эффекторной клетке, так как пресинаптическое окончание чувствительно только к нервному импульсу, а постсинаптическая мембрана - к медиатору.
Неизолированное - возбуждение рядом расположенных постсинаптических мембран суммируется.
Синаптическая задержка в передаче сигнала к другой клетке (в нервно-мышечном синапсе 0,5-1,0 мс), что связано с высвобождением медиатора из нервного окончания диффузией его к постсинаптической мембране и возникновением постсинаптических потенциалов, способных вызвать ПД.
Декрементность (затухание ) возбуждения в химических синапсах при недостаточном выделении медиатора из пресинаптических окончаний в синаптические щели.
Низкая лабильность (в нервно-мышечном синапсе составляет 100 Гц), которая в 4 - 8 раз ниже лабильности нервного волокна. Это объясняется синаптической задержкой.
Проводимость нервно-мышечного синапса (как и химических синапсов ЦНС) угнетается или, наоборот, стимулируется различными веществами .
Например, кураре и курареподобные вещества (диплацин, тубокурарин) обратимо связываются с N-холинорецепторами постсинаптической мембраны, блокируют действие на нее ацетилхолина и передачу в синапсе. Напротив, некоторые фармакологические препараты, например прозерин, подавляют активность ацетилхолинэстеразы, способствуя умеренному накоплению ацетилхолина и облегчению синаптической передачи, что используется в лечебной практике.
Утомляемость (синаптическая депрессия) - ухудшение проводимости вплоть до полной блокады проведения возбуждения при длительном функционировании синапса (главная причина - истощение медиатора в пресинаптическом окончании).
Вопросы для самоконтроля
1.Каков механизм распространения возбуждения по нервному волокну? Какова роль перехватов Ранвье в проведении возбуждения по миелинизированному нервному волокну?
2.В чем преимущество скачкообразного (сальтаторного) распространения возбуждения над непрерывным его проведением вдоль мембраны волокна?
3.В чем физиологическое значение изолированного проведения возбуждения по нервному волокну?
4.Какие нервные волокна (афферентные или эфферентные, вегетативные или соматические) относятся к группе А? Какова скорость проведения возбуждения по ним?
5.Какие нервные волокна (афферентные или эфферентные, вегетативные или соматические) относятся к группе В? Какова скорость проведения по ним?
6.Какие нервные волокна (афферентные или эфферентные, вегетативные или соматические) относятся к группе С? Какова скорость проведения возбуждения по ним?
7.Перечислите структуры нервно-мышечного синапса (скелетная мышца). Что называют концевой пластинкой?
8.Перечислите последовательность процессов, ведущих к освобождению медиатора из пресинаптической мембраны в синаптическую щель при передаче возбуждения в синапсе.
9. Локальным потенциалом или распространяющимся возбуждением является потенциал концевой пластинки?
10.Что такое миниатюрные потенциалы концевой пластинки, каков механизм их возникновения?
11.В чем заключается трофическое влияние нерва на мышцу, осуществляемое через нервно-мышечный синапс?
12.Какие вещества являются медиаторами в нервно-мышечных синапсах гладкой и поперечнополосатой мышц?
13.Что такое сенсорный рецептор?
14.На какие две группы делятся сенсорные рецепторы по скорости адаптации? Назовите рецепторы, относящиеся к каждой из них.
15.Что понимают под первичными и вторичными рецепторами?
16.Перечислите основные свойства рецепторов.
17.Что называют адаптацией рецепторов? Как изменяется частота импульсов в афферентном нервном волокне при адаптации рецептора?
18.Назовите локальные потенциалы, возникающие при возбуждении первичных и вторичных рецепторов.
19.Рецепторный потенциал, где он возникает, каково его значение?
20.Генераторный потенциал, где он возникает, каково его значение?
21.Где возникает потенциал действия при возбуждении первичного сенсорного рецептора?
22. Где возникает потенциал действия при возбуждении вторичного сенсорного рецептора?
Физиология мышц
1.3.1. Структурнофункциональная характеристика скелетной мышцы
Мышцы подразделяют на поперечнополосатые (скелетная и сердечная ) и гладкие (сосуды и внутренние органы, кроме сердца).
Скелетная мышца состоит из мышечных волокон , изолированных в структурном и функциональном отношении друг от Друга, которые представляют собой вытянутые многоядерные клетки. Толщина волокна составляет 10-100 мкм, а его длина варьирует в пределах от нескольких миллиметров до нескольких сантиметров. Количество мышечных волокон, установившись постоянным на 4-5-м месяце постнатального онтогенеза, в последующем не изменяется; с возрастом изменяются (увеличиваются) лишь их длина и диаметр.
Назначение основных структурных элементов. Характеристика основных элементов мышечного волокна. От клеточной мембраны мышечного волокна (сарколеммы) вглубь отходят многочисленные поперечные инвагинации (Т-трубочки ), которые обеспечивают ее взаимодействие с саркоплазматическим ретикулулом (СПР ) (рис. 20).

Рис. 20. Взаимоотношение клеточной мембраны (1), поперечных трубочек (2), боковых цистерн (3) и продольных трубочек (4) саркоплпзматическаого ретикулума, сократительных белков (5): А – в состоянии покоя; Б – при сокращении мышечного волокна; точками обозначены ионы Ca 2+
СПР представляет собой систему связанных друг с другом цистерн и отходящих от них в продольном направлении канальцев, расположенных между миофибриллами. Терминальные (концевые) цистерны СПР примыкают к Т-трубочкам, формируя так называемые триады . В цистернах содержится Са 2+ , играющий важную роль в мышечном сокращении. В саркоплазме имеются внутриклеточные элементы: ядра, митохондрии, белки (в том числе миоглобин), капельки жира, гранулы гликогена, фосфатсодержащие вещества, различные малые молекулы и электролиты.
Миоибриллы - субъединицы мышечного волокна. В одном мышечном волокне может насчитываться более 2 тыс. миофибрилл, их диаметр 1-2 мкм. В одиночной миофибрилле содержится 2-2,5 тыс. протофибрилл - параллельно расположенных нитей белка (тонкие - актин, толстые - миозин ). Актиновые нити состоят из двух субъединиц, скрученных в виде спирали. В состав тонких нитей входят также регуляторные белки - тропомиозин и тропонин (рис. 21).
Рис. 21. Взаимное расположение структурных элементов миофибрилл при их расслаблении (А,Б) и сокращении (В)
Эти белки в невозбужденной мышце препятствуют взаимосвязи актина и миозина, поэтому мышца в покое находится в расслабленном состоянии. Миофибриллы включают в себя последовательно соединенные блоки - саркомеры (Б), отделенные друг от друга Z-полосками. Саркомер (длина 2-Змкм) является сократительной единицей мышечного волокна; при длине 5см оно включает в себя около 20 тыс. последовательно соединенных саркомеров. Миофибриллы отдельного мышечного волокна связаны таким образом, что расположение саркомеров совпадает, и это создает картину поперечной исчерченности волокна при наблюдении в световом микроскопе (рис. 22).
Рис. 22. Саркомер миоцита скелетной мышцы (A. Vander, J. Sherman, D. Luciano, 2004)
Элементы саркoмера (см. рис. 21). Миозиновые протофибриллы образуют наиболее темную часть саркомера - А-диск (анизотропный, он сильно поляризует белый свет). Более светлый участок в центре А-диска называют Н-зоной . Светлый участок саркомера между двумя А-дисками называют 1-диском (изотропный, почти не поляризует свет). Он образован актиновыми протофибриллами, идущими в обе стороны от Z-полосок. Каждый саркомер имеет два набора тонких нитей, прикрепленных к Z-полоскам, и один комплект толстых нитей, сосредоточенных в А-диске. В расслабленной мышце концы толстых и тонких филаментов в разной степени перекрывают друг друга на границе между А- и 1-дисками.
Классификация мышечных волокон:
По структурно-функциональным свойствам и цвету выделяют две основные группы мышечных волокон: быстрые и медленные.
Белые (быстрые) мышечные волокна содержат больше миофибрилл и меньше - митохондрий, миоглобина и жиров, но больше гликогена и гликолитических ферментов; эти волокна называют гликолитическими . Капиллярная сеть, окружающая эти волокна, относительно редкая. Скорость рабочего цикла у данных волокон примерно в 4 раза больше, чем у медленных, что объясняется более высокой АТФазной активностью быстрых волокон, но они обладают малой выносливостью. У белых мышечных волокон число нитей актина и миозина больше, чем у красных, поэтому они толще и сила их сокращения больше, чем у красных волокон.
Красные мышечные волокна содержат много митохондрий, миоглобина , жирных кислот. Эти волокна окружены густой сетью кровеносных капилляров, они имеют меньший диаметр. Митохондрии обеспечивают высокий уровень окислительного фосфорилирования, поэтому данные волокна называют оксидативными. Красные мышечные волокна подразделяются на две подгруппы: быстрые и медленные . Медленные волокна могут выполнять работу в течение относительно продолжительного периода времени; утомление в них развивается медленнее. Они более приспособлены к тоническим сокращениям. Красные быстрые волокна по скорости утомления занимают промежуточное положение между белыми и красными медленными. Скорость их сокращения близка к скорости сокращения белых волокон, что также объясняется высокой АТФазной активностью миозина красных быстрых волокон.
Также имеется незначительное число истинных тонических мышечных волокон; на них локализуется по 7-10 синапсов, принадлежащих, как правило, нескольким мотонейронам, например, в глазодвигательных мышцах, мышцах среднего уха. ПКП этих мышечных волокон не вызывают генерации ПД в них, а непосредственно запускают мышечное сокращение.
Группа мышечных волокон, двигательную (нейромоторную) единицу. В мышцах, совершающих быстрые и точные движения, например в глазодвигательных, нейромоторные единицы состоят из 3-5 мышечных волокон. В мышцах, осуществляющих менее точные движения (например, мышцы туловища и конечностей), двигательные единицы включают сотни и тысячи мышечных волокон. Большая двигательная единица, по сравнению с малой, включает крупный мотонейрон с относительно толстым аксоном, который образует большое число концевых веточек в мышце и, следовательно, иннервирует большое число мышечных волокон. Все мышечные волокна одной двигательной единицы, независимо от их количества, относятся к одному типу. Все скелетные мышцы по своему составу являются смешанными, т.е. образованы красными и белыми мышечными волокнами.
Специфическим свойством всех мышц является сократимость - способность сокращаться, т.е. укорачиваться или развивать напряжение. Реализация этой способности осуществляется с помощью возбуждения и его проведения по мышечному волокну (свойства соответственно возбудимости и проводимости).
Скелетные мышцы не обладают автоматией, управляются организмом произвольно импульсацией из ЦНС, поэтому их называют также произвольными . Гладкие мышцы по собственному желанию не сокращаются, поэтому их называют также непроизвольными, но они обладают автоматией.
Функции скелетной мышцы :
Обеспечение двигательной активности организма - поиск и добывание воды и пищи, ее захват, жевание, глотание, оборонительные реакции, трудовая деятельность - физическая и творческая работа художника, писателя, ученого, композитора в конечном итоге выражается в движении: рисование, письмо, игра на музыкальном инструменте и т.п.
Обеспечение дыхания (движений грудной клетки и диафрагмы).
Коммуникативная функция (устная и письменная речь, мимика и жесты).
Участие в процессах терморегуляции организма с помощью изменения интенсивности сократительного термогенеза.
©2015-2019 сайт
Все права принадлежать их авторам. Данный сайт не претендует на авторства, а предоставляет бесплатное использование.
Дата создания страницы: 2016-04-15
Нервное волокно непосредственно не контактирует с мышечным. Между ними очень узкая щель (20-50 нм). Нервное окончание, часть мышечного волокна, к которой прилегает это окончание, и щель между ними составляют нервно-мышечный синапс. Нервное волокно заканчивается синаптической бляшкой, в которой содержатся Мтохондрии и значительное количество (около 300 тыс.) пузырьков, где концентрируется медиатор - вещество, с помощью которой возбуждение распространяется с нерва на мышцу. В нервно-мышечном синапсе этим медиатором является ацетилхолин. Синтез ацетилхолина происходит в синаптической бляшке и требует энергетических затрат. Часть бляшки, прилегающей к щели, называется пресинаптической мембраной. С другой стороны щели расположен постсинаптическая мембрана, которая принадлежит мышечному волокну. Часть этой мембраны, которая непосредственно прилегает к бляшки, называется конечной пластинкой.
В состав постсинаптической мембраны входят рецепторные белки (молекулярная масса - около 275 000), которые реагируют на ацетилхолин и называются холинорецепторами. Они реагируют также на никотин, отсюда их название - Н-холинорецепторы. Реакция ацетилхолина с Н-холинорецепторы приводит конформационные изменения молекулы рецептора. Это влияет на рядом расположенный хемочутливий ионный канал, который может пропускать Na +, К +, Са2 +. Белковые структуры этих каналов имеют отрицательный заряд, и поэтому анионы через них не проходят.
Передача информации через нервно-мышечный синапс происходит в такой последовательности:
1. Нервным волокном в бляшки приходит ПД.
2. Вследствие действия ПД на мембрану нервного окончания открываются Са2 +-каналы и эти ионы вступают в бляшку.
3. Повышение в бляшке концентрации Са2 + приводит (при участии кальмодулина) до выхода медиатора из пузырьков в синаптическую щель. Вследствие действия единичного ПД медиатор выходит примерно из 300 пузырьков.
4. Ацетилхолин диффундирует через щель.
5. Ацетилхолин реагирует с Н-холинорецепторы.
6. Открываются «ворота» хемочутливих каналов и за концентрационным градиентом Na + переходит в мышечное волокно, а К + - наружу.
7. Происходит деполяризация концевой пластинки и развивается ее потенциал (ПКП). ПКП-один из видов местного возбуждения. Чем больше выделяется ацетилхолина, то в большей степени выражен этот потенциал.
8. Когда ЛКП достигает критической величины (ЕКС), деполяризуется колосинаптична електрозбудлива мембрана и развивается
ПД мышечного волокна, которое распространяется по обе стороны синапса. В этом процессе участвуют потенциалзависимые ионные каналы.
Ацетилхолин выделяется в незначительном количестве даже тогда, когда ПД нервными волокнами не поступает. Даже в таком количестве (кванты медиатора) он обуславливает незначительную деполяризацию постсинаптической мембраны (доли мВ), которая имеет название миниатюрных потенциалов концевой пластинки (МПКП).
Ацетилхолин, выделившегося в синаптическую щель, очень быстро разрушается ферментом ацетилхолинэстеразой. Вследствие этого образуется холин, который попадает в синаптическую бляшку и участвует в создании новых порций ацетилхолина.
Передача информации в нервно-мышечном синапсе происходит в одном направлении, с определенной задержкой (около 0,5 мс). При слишком частых и длительных раздражениях могут наступать истощение запасов ацетилхолина и усталость, являются методы усиления или ослабления нередачи импульсов нервно-мышечных синапсов. Так, усилить передачу их можно с помощью заменителей ацетилхолина-холиномиметическим средств (например, карбахолин) или путем инактивации ацетилхолинэстеразы (езерин, физостигмин). В этом случае медиатор не разрушается, его действие усиливается и продлевается.
Ослабить или прекратить передачу можно при введении курареподобных веществ - миорелаксантов (например тубокурарина, диплацина). Эти препараты связываются с холинорецепторами и блокируют действие ацетилхолина.
Физиология нервов.
Центральная нервная система у человека состоит из нервных клеток, каждая из которых имеет один аксон и много дендритов. Нервные волокна делятся на: мякотные и безмякотные. Все они имеют шванновскую оболочку, а мякотные, кроме этого, покрыты еще миелиновой оболочкой между которой имеются перехваты Ранвье, в которых миелиновая оболочка отсутствует. Безмякотные волокна имееют малый диаметр, меньше 1,3 мкм, небольшую скорость распространения импульса до 2 м/сек, продолжительность ПД – 2 мсек. Мякотные возникли из безмякотных, диаметр до 25 мкм, скорость распространения возбуждения до 120 м/сек, продолжительность ПД – 0,4-0,5 мсек.
Распространение ПД по безмякотному волокну:
немецкий физиолог Герман предложил теорию «местных токов», согласно которой при распространении возбуждения на мембране нервного волокна между возбужденным и невозбужденным участком возникает местный ток, который является раздражителем для невозбужденного участка. Если его величина достаточна для возникновения ПД в соседнем участке, то ПД распространяется на этот участок.
Распространение местных токов в безмякотных нервных волокнах прямопропорционально сопротивлению мембраны и обратнопропорционально сопротивлению внутри- и внеклеточной среды. Расстояние на которое распространяются местные токи будет тем больше, чем больше амплитуда ПД и чем меньше пороговый потенциал.
В мякотных волокнах возбуждение возникает при нанесении раздражения в перехватах Ранвье (теория Тасаки) и распространяется по мембране волокна сальтаторно (скачкообразно) (рис.1).
Рисунок 1. Проведение возбуждения по миелиновому нервному волокну.
При этом может охватывать не только один, но и два перехвата Ранвье, что обеспечивает надежность, а также увеличивает скорость распространения возбуждения и экономически более выгодна, так как на 1 импульс в безмякотном волокне энергия расходуется в 20 раз больше, чем в мякотном. Таким образом, скорость и расстояние, на которое распространяется возбуждение в безмякотных волокнах пропорциональна диаметру, сопротивлению мембраны и амплитуде ПД; в мякотных прямопропорциональна длине межперехватных участков, а их длина тем больше, чем больше диаметр волокна. Скорость не зависит от силы раздражения.
Теория Германа была экспериментально подтверждена.
Гельмгольц – определил скорость распространения импульса у лягушки; Бабский – определил скорость распространения импульса у человека. Эрландер и Гассер классифицировали различные нервные волокна на три группы А, В и С:
Типы нервных волокон
А – миелинизированные, наибольшего диаметра, скорость 120-70 м в сек, длительность ПД – минимальная, делятся на подгруппы: альфа, бета, гамма, дельта. Пример – аксоны мотонейронов.
В – миелинизированные волокна, меньшего диаметра, скорость 3-18 м в сек, ПД более длителен. Пример – преганглионарные волокна симпатической нервной системы.
С – немиелизированные нервные волокна, скорость менее 2 м в сек, длительность ПД наибольшая. Пример - постганглионарные волокна парасимпатической нервной системы.
Законы проведение возбуждения по нервам.
I закон
анатомо-физиологический целостности нервного волокна
. Чтобы возбуждение распространялось по нервному волокну необходимо не только его морфологическая целостность, но и физиологическая непрерывность. Препараты для проводниковой анестезии нарушают физиологическую непрерывность тем, что инактивируют натриевую проницаемость в нервных волокнах.
II закон
изолированного проведения возбуждения по нервному волокну
. В смешанном нерве возбуждение с одного нервного волокна не передается на соседние, так как сопротивление межклеточной жидкости меньше чем сопротивление мембран соседних волокон. Этим обеспечив
ается точность проведения информации в нервных волокнах к иннервируемым структурам.
III закон
двухстороннее проведение возбуждения
. Распространение ПД по мембране нервного волокна возможно в обе стороны, так как строение мембраны на всем протяжении одинаково. В то же время возбуждение не может возвратиться в участок, где оно возникло, так как он находится в состоянии рефрактерности.
Парабиоз.
Н.Е. Введенский, исследуя прохождение импульса через отрезок нерва на который воздействую химические или наркотические вещества (альтераторы), наблюдал резкое снижение лабильности. Парабиоз характеризуется постепенным развитием, в котором можно выделить четыре фазы:
I Продромальная (не всегда проявляется, так как очень кратковременная) характеризуется: повышением возбудимости, повышением лабильности.
II Уравнительная – эффекты от сильных и слабых раздражителей уравновешиваются.
III Парадоксальная - на сильные, либо частые раздражения эффект бывает меньше, чем на слабые или редкие.
IV Тормозная – ни сильные, ни слабые раздражения не вызывают сокращения мышц. Через поражённый участок не проходят импульсы.
Если второй парой электродов подействовать на поражённый участок, то возбуждение будет, т.е. ткань ещё жива.
Если снять альтератор, то ткань возвращается к исходному состоянию в обратном порядке фаз IV, III, II, I.
Парабиоз – это стойкое не распространяющееся возбуждение.
Возникают потенциалы меньшие по своей амплитуде, а дальше абортивные потенциалы, не способные распространяться: уменьшаются процессы Na-евой проницаемости, и увеличиваются процессы Na-евой инактивации.
Нервно-мышечная передача.
Взаимодействие человека с внешней средой не возможно представить без его мышечной системы. Производимые движения скелетной мускулатурой необходимы как для выполнения простейших перемещений тела в пространстве, сложных манипуляций хирурга, стоматолога, выражения самых тонких чувств и мыслей с помощью речи, мимики, жестов. Работа сердца обеспечивает кровоснабжение всех органов, работа гладких мышц создает условия для нормального осуществления физиологических процессов, обеспечивающих гомеостаз, практически во всех системах: гастроинтестинальной, сердечно-сосудистой, выделительной, репродуктивной, дыхательной. Ведущая роль скелетной мускулатуры также в производстве тепла и поддержании температуры тела. Мышцы - это «машины», преобразующие химическую энергию в механическую (работу) и тепло. Масса мышц больше чем других органов, 40-50% от массы тела.
В естественных условиях (в нашем организме) возбуждение мышечного волокна (или нескольких мышечных волокон, составляющих мышцу) возникает в результате передачи возбуждения с нервного волокна на мембрану мышечного в местах контакта нерва и мышцы: нервно-мышечных синапсах.
Механизм нервно-мышечной передачи
Синапсы представляют собой коммуникационные структуры, которые формируются окончанием нервного волокна и прилегающей к нему мембраной мышечного волокна (пресинаптической нервной и постсинаптической мышечной мембранами) (рис.2). 
Рис.2. Мионевральный синапс.
Когда нервный импульс достигает окончания аксона, на деполяризованной пресинаптической мембране открываются потенциалзависимые Са2+ каналы. Вход Са2+ в аксональное расширение (пресинаптическую мембрану) способствует высвобождению химических нейромедиаторов, находящихся в виде везикул (пузырьков) из окончания аксона. Медиаторы (в нервно-мышечном синапсе это всегда ацетилхолин) синтезируются в соме нервной клетки и путем аксонального транспорта транспортируются к окончанию аксона, где и выполняют свою роль. Медиатор диффундирует через синаптическую щель и связывается со специфическими рецепторами на постсинаптической мембране. Так как медиатором в нервно-мышечном синапсе является ацетилхолин, то рецепторы постсинаптической мембраны называют холинорецепторами. В результате этого процесса на постсинаптической мембране открываются хемочувствительные Nа+-каналы, возникает деполяризация, величина которой различна, и зависит от количества выделенного медиатора. Чаще всего возникает локальный процесс, который называют потенциалом концевой пластинки (ПКП). При повышении частоты стимуляции нервного волокна, усиливается деполяризация пресинаптической мембраны, а, следовательно, возрастает количество выделяемого медиатора и число активированных хемочувствительных Nа+каналов на постсинаптической мембране. Таким образом, возникают ПКП, которые по амплитуде деполяризации суммируются до порогового уровня, после чего, на мембране мышечного волокна, окружающей синапс, возникает ПД, который обладает способностью к распространению вдоль мембраны мышечного волокна. Чувствительность постсинаптической мембраны регулируется активностью фермента – ацетилхолинэстеразы (АЦХ-Э), который гидролизует медиатор АЦХ на составные компоненты (ацетил и холин) и возвращает назад – в пресинаптическую бляшку для ресинтеза. Без удаления медиатора на постсинаптической мембране развивается длительная деполяризация, которая ведет к нарушению проведения возбуждения в синапсе – синаптической депрессии. Таким образом, синаптическая связь обеспечивает одностороннее проведение возбуждения с нерва на мышцу, однако на все эти процессы расходуется время (синаптичекая задержка), что приводит к низкой лабильности синапса по сравнению с нервным волокном.
Таким образом, нервно-мышечный синапс
является «выгодным» местом,
куда можно воздействовать фармакологическими препаратами, изменяя чувствительность рецептора, активность фермента. Эти явления будут часто встречаться в практике врача: например, при отравлении токсином ботулизма – блокируется высвобождение медиатора АЦХ (разглаживание морщин в косметической медицине), блокада холиноререпторов (курареподобными препаратами, бунгаротоксином) нарушает открытие Nа+ каналов на постсинаптической мембране. Фосфоорганические соединения (множество инсектицидов) нарушает эффективность АЦХ-Э и вызывает длительную деполяризацию постсинаптической мембраны. В клинике используют специфические блокаторы нервно-мышечного проведения: блокада холинорецепторов курареподобными препаратами, сукцинилхолином и другими конкурентными ингибиторами, вытесняющими АЦХ с холинорецептора. При заболевании миастении из-за дефицита холинорецепторов на постсинаптической мембране (из-за их аутолитического разрушения) возникает прогрессирующая мышечной слабость, вплоть до полной остановки мышечных сокращений (остановка дыхания). В этом случае используют блокаторы АЦХ-Э, что приводит к увеличению длительности связывания медиатора с меньшим количеством холинорецепторов и несколько увеличивает амплитуду деполяризации постсинаптической мембраны.
Физиология мышц.
Существует 3 типа мышечной ткани: поперечно-полосатая, сердечная, гладкая.
Скелетная
мышечная ткань образует большой объем соматической мускулатуры. Содержит хорошо выраженное упорядоченное строение сократительных белков в виде поперечной исчерченности. Связей между отдельными мышечными волокнами нет. Регуляция сокращений происходит сознательно.
Сердечная
мышца
содержит также поперечную исчерченность, но является функциональным синцитием. Благодаря наличию пейсмекерных клеток, обладает способностью генерировать спонтанные нервные импульсы, обеспечивающие сердечные сокращения.
В гладких
мышцах нет поперечной исчерченности, которая бы придавала определенные физические и физиологические свойства этой ткани. Так в отличие от скелетной, которой присуща высокая эластичность, в гладкой мышце больше выражено свойство пластичности, что обусловлено отсутствием четкой упорядоченности миофиламентов актина и миозина. В отличие от регулярной саркомерной структуры скелетных и сердечной мышц, тонкие филаменты гладких мышц присоединены к структурам в цитоплазме, называемым плотными тельцами
(прикрепительными бляшками сарколеммы), состоящими из белка десмина
. Гладкие мышцы бывают висцеральные (мультиунитарные) и полиэлементные (унитарные).
Висцеральные
содержат мостики- щелевые контакты с низким сопротивлением электрическому току – синцитий. Висцеральные мышцы встречаются в стенках полых органов (кишечник, матка, мочеточники, мочевой пузырь).
Полиэлементные
гладкие мышцы состоят из отдельных мышечных единиц без соединительных мостиков, поэтому для них характерны точные, четко дозированные сокращения, подобно скелетным. Однако, сокращения этих мышц нельзя контролировать, в отличие от скелетных (мышцы радужки глаза, цилиарного тела, семенных протоков, артерии). Благодаря своему строению (висцеральные)
обладают большой пластичностью, однако при определенной предельной степени растяжения способны деполяризоваться и сокращаться (саморегуляция). Пример, давление внутри стенок мочевого пузыря мало изменяется при относительно небольшом его растяжении, если растяжение возрастает резко – происходит сокращение мышц детрузора – эвакуация мочи даже в тех случаях, когда иннервация нарушена. Полиэлементные
гладкие мышцы имеют более мощную (плотную) иннервацию и слабое развитие межклеточных контактов (нексусов). Тонус этих мышц и его колебания имеют нейрогенную природу. Имеют парасимпатические и симпатические, а также метасимпатические волокна. Строение нервных окончаний в гладкой мышце отличается от строения нервно-мышечного синапса в скелетной мышцы. В гладкой мышце нет концевых пластинок и отдельных нервных окончаний. По всей длине разветвлений адренергических и холинергических нервных волокон имеются утолщения, называемые варикозами. Они содержат гранулы с медиатором, который выделяется из каждой варикозы. Клетки, лишенные непосредственных контактов с варикозами, активируются ПД, распространяющимися через нексусы на соседние клетки. По ходу следования нервного волокна мышечные клетки могут возбуждаться или тормозиться (стимуляция адренергических волокон уменьшает, а холинергических – увеличивает мышечную активность, в других, например в сосудах, норадреналин – усиливает, а ацетилхолин – уменьшает мышечный тонус. Ионная природа гладкой мышцы определяется особенностями каналов мембраны гладко-мышечной клетки. Основную роль в механизме генерации ПД играют ионы Са2+, но по этим каналам могут внутрь клетки двигаться и другие двухвалетные ионы Ва2+, Mg2+. Вход Са2+ в клетку необходим для поддержания тонуса мышц и развития сокращения, поэтому блокирование Са2+ каналов гладких мышц приводит к ограничению поступления этого иона в цитоплазму миоцитов внутренних органов и сосудов, что широко используется в практической медицине для коррекции моторной функции ЖКТ и тонуса сосудов.
Регуляция мышечных сокращений зависит от объема иннервации. Аксон спинно-мозгового (двигательного) мотонейрона ветвится на несколько терминалей (веточек), каждая из которых подходит к одному мышечному волокну. Поэтому в целостном организме, при возбуждении 1 нервного волокна сокращается группа мышц. Совокупность двигательного нейрона и мышечных волокон, которые он иннервирует называют двигательной единицей. Количество мышечных волокон, входящих в двигательную единицу, различно и зависит о функции, которую реализует каждая конкретная мышца в организме. В мышцах глаз, кисти руки, которые обеспечивают высокоточные, координированные движения, 1 двигательная единица содержит 3-5 мышечных волокон. Двигательные единицы мышц спины, бедра – состоят из нескольких сотен мышечных волокон, которые регулируются одним мотонейроном.
Вернемся к мембране мышечного волокна, на которой распространяется ПД, возникший в результате «успешного» проведения возбуждения через синапс. Продолжением мембраны мышечного волокна является саркотубулярная система, образующая поперечные инвагинации (впячивания) (Т-система поперечных трубочек). Система Т-трубочек выполняет несколько важных функций: является внутриклеточным депо ионов Са2+; сообщается с внеклеточной жидкостью и таким образом регулируется содержание Са2+ в ней; содержит потенциалчувствительные Nа+ каналы, которые дают возможность ПД распространяться как вдоль, так и вглубь мышечного волокна. Система Т-трубочек обеспечивает быстрое и согласованное возбуждение мышечной клетки, так как распространение деполяризации по Т-системе сопряжено с выходом Са2+ из цистерн саркоплазматического ретикулума, обеспечивает инициацию сокращения мышцы. Через Т-трубочки может происходить выделение продуктов обмена (например, молочной кислоты) из мышечной клетки в интерстиций (межклеточное пространство) и далее в кровь. Большой объем клетки скелетной мышцы невозможно было бы активировать быстро, если бы Са2+ поступал из внеклеточной среды. Накоплению Са2+ в саркоплазматическом ретикулуме способствует белок (кальсеквестрин), который непрочно связывает Са2+ в ретикулуме. Высвобождение Са2+ из саркоплазматического ретикулума в скелетных мышцах происходит благодаря взаимодействию Т трубочек и концевых цистерн (триада).
Ионы Са2+ попав в саркоплазму инициируют сокращение, связываясь с белком тропонином – блокатором актина в покое (рис.3).

Рисунок. 3. Механизм мышечного сокращения.
Тонкая нить актина состоит из 2 тяжей фибриллярного актина, на котором нанизаны отдельные глобулы мономера актина (как бусы). Актин содержит активные центры (сайты) связывания с миозином, которые в состоянии покоя заблокированы тропонином. Тропонии подавляет АТФ-азную активность миозина, что делает невозможным расщепление АТФ и мышечные волокна пребывают в расслабленном состоянии. Крупный сократительный белок – миозин, состоящий из 6 полипептидных цепей, уложенных попарно. 2 из них - тяжелые цепи миоглобина, обладающие свойством ферментов. Связанный с трононином Са2+ освобождает активные центры актина для контакта с миозином. В присутствии актина миозиновый фермент (глобулярная головка) разрушает АТФ и взаимодействует с тонкими нитями актина, создавая движущую силу сокращения – образуя поперечные мостики («гребки») и мышца укорачивается (сокращается). Таким образом, АТФ обеспечивает сокращение энергией, гидролизуясь на глобулярных головках миозина. Энергия (Э) которая высвобождается при гидролизе АТФ, превращается в силу сокращения за счет конформационных (пространственных) изменений в миозине (рабочий ход - образование поперечных гребковых мостиков) – это тепло активации, выделяющее при связывании актина и миозина. АДФ, связанная с миозином, уменьшает сродство поперечного мостика к активному центру актина, что инициирует следующую фазу – мышечное расслабление. За счет гидролиза АТФ выделяется Э (тепло укорочения), которая идет на:
1) работу Са-АТФ-азы, (активация насоса происходит за счет неорганического фосфата, образующегося при гидролизе АТФ),
2) за счет Э происходит откачивание против градиента концентрации Са2+ назад в саркоплазматический ретикулум (активный транспорт).
3) АТФ в мышечной клетке обеспечивает работу Na+-К-АТФ-азы, обеспечивающей удаление Na+ из клетки и восстановление потенциала покоя (а, следовательно, и возбудимости) мышечной клетки.
АТФ обеспечивает процессы, как сокращения, так и расслабления. Если Са2+ не будет транспортироваться назад в СПР, то расслабление не наступит, развивается ригидность мышцы (при трупном окоченении), или в живом организме – при посттетанической контрактуре – накопление Са2+ в саркоплазме инициирует длительное мышечное сокращение вне поступления ПД через синапс на мембрану мышечного волокна.
Гладкая мышца содержит также тропомиозин, но не имеет тропонина, соотношение актина к миозину 14-16 к 1, сравните в скелетных соотношение актина к миозину 2 к 1. Гладкая мышца имеет щелевые контакты – это мостики, соединяющие мембраны соседних клеток. Регуляция сократительной активности гладких мышц происходит благодаря связыванию Са2+ с кальмодулином, активирующим киназу легкой цепи миозина, которая приводит к гидролизу АТФ и запускает цикл образования поперечных мостиков.
ПД скелетной мышцы длится около 2-4- мс и проходит по мембране мышечного волокна со скоростью около 5 м/с. 1 ПД вызывает одиночное мышечное сокращение, которое начинается через 2 мс после начала деполяризации мембраны (латентный период) и завершается сокращение почти одновременно с реполяризацией. Длительность одиночного сокращения различна и зависит от типа мышечной ткани. При частых стимулах развивается суммарное мышечное сокращение всех мышечных волокон, обладающих различным сопротивлением мембран к электрическому току. Однако, незначительные отличия порогов возбуждения обеспечивают синхронность мышечного сокращения целой мышцы. Наличие абсолютного рефрактерного периода около 1-3 мс, обуславливает возникновение различных видов тетанусов (суммарных мышечных сокращений). Существует зубчатый и гладкий тетанусы. Частота стимуляции мышцы для развития гладкого тетануса должна быть выше, чем для развития зубчатого. Стимул должен попадать в фазу укорочения мышцы, если же мышцы начала расслабляться, а мы ее стимулирует, то получаем – зубчатый тетанус. Минимальный промежуток времени между последовательными эффективными стимулами во время тетануса не может быть меньше рефрактерного периода, которые приблизительно соответствует длительности ПД. Поскольку мышцы состоят из мышечных волокон с различным уровнем возбудимости, имеется определенная зависимость между величиной стимула и ответной реакцией. Увеличение силы сокращения возможно до определенного предела, после которого амплитуда сокращения остается неизменной при увеличении амплитуды стимула (надо отметить, то в мышце суммируются не ПД, а сокращения). При этом все волокна, входящие в состав мышцы принимают участие в сокращении.
В организме человека имеются быстрые, фазные мышечные волокна (белые), длительность сокращения которых до 7,5 мс, и медленные, тонические (красные), которые обеспечивают сильные и мощные движения, длящиеся до 100 мс. Красные (тонические) имеют много волокон миозина типа I, которые отличаются низкой активностью АТФ-азы миозина. Скорость расщепления АТФ является фактором, предопределяющим частоту гребковых движений, и таким образом, скорость скольжения нитей актина вдоль миозина. Из СПР Са2+ транспортируется медленно, высокая окислительная способность, много капилляров, много миоглобина в структуре миозина (тяжелые цепи), много митохондрий. На стимул реагируют медленно, имеют длительный латентный период сокращения, поэтому способны к длительным, медленным, тоническим сокращениям, более резистентны к утомлению. Главная функция – поддержание положения тела. Белые мышцы содержат волокна миозина II типа. Гликолитический тип окисления, мало миоглобина, митохондрий, это волокна большого диаметра с высокой активность АТФ-азы миозина, способны развить значительную силу, но быстро утомляются.
Сила мышц определяется тем максимальным грузом, который мышца в состоянии чуть-чуть приподнять. Сила различных мышц неодинакова. Для сравнения силы разных мышц максимальный груз, который мышца в состоянии поднять делят на число квадратных сантиметров ее физиологического поперечного сечения. Силовые характеристики выше у мышц с перистым (косым) расположением волокон, при этом физиологическое сечение больше геометрического поперечного сечения. Сумма поперечного сечения не всегда совпадает с физиологическим сечением мышцы (только при параллельном расположении волокон.
Сила сокращения
изолированной скелетной мышцы при прочих равных условиях зависит от исходной длины мышцы. Умеренное растяжение мышцы приводит к тому, что развиваемая ею сила возрастает по сравнению с силой, которую развивает нерастянутая мышца. Происходит суммирование пассивного напряжения, обусловленного наличием эластических компонентов мышцы, и активного сокращения (переход к правилу средних нагрузок – физиологический механизм данного закона). Способность совершать работу
определяется произведением величины поднятого груза на высоту подъема. Величина работы мышцы постепенно увеличивается с увеличением массы поднимаемого груза, но до определенного предела, после которого увеличение массы груза приводит к снижению величины работы, так как высота подъема груза резко падает. Следовательно, максимальная работа совершается мышцей при средних величинах нагрузок.
Сила сокращения и работа не остаются постоянными при статической и динамической работе. В результате продолжительной деятельности работоспособность скелетной мускулатуры понижается. Это явление называют утомлением.
При этом снижается сила сокращений, увеличивается латентный период сокращения и период расслабления. Статический режим работы более утомителен, чем динамический (почему объяснить). Накапливаются продукты процессов окисления- молочная пировиноградная кислота, которые снижают возможность генерирования ПД. Нарушаются процессы ресинтеза АТФ и креатинфосфата, необходимых для энергетического обеспечения мышечного сокращения. В естественных условиях мышечное утомление при статической работе в основном определяется неадекватным регионарным кровотоком. Возникает «кислородное голодание» и утомление прогрессивно нарастает.
В 1903 году Сеченов – восстановление работоспособности утомленной мышцы значительно ускоряется при совершении работы другой мышцей в период отдыха утомленной – такой отдых называют активным.
Однако высокий уровень умственной деятельности ускоряет процессы развития утомления в мышечной системе (утомление нервных центров).