Rakumembraani põhifunktsioon. rakumembraan.
Kõik elusorganismid Maal koosnevad rakkudest ja iga rakku ümbritseb kaitsekesta – membraan. Kuid membraani funktsioonid ei piirdu ainult organellide kaitsmise ja ühe raku teisest eraldamisega. Rakumembraan on keeruline mehhanism, mis on otseselt seotud paljunemise, regeneratsiooni, toitumise, hingamise ja paljude teiste oluliste rakufunktsioonidega.
Mõistet "rakumembraan" on kasutatud umbes sada aastat. Sõna "membraan" tähendab ladina keelest "filmi". Aga rakumembraani puhul oleks õigem rääkida kahe teatud viisil omavahel ühendatud kile kombinatsioonist, pealegi on nende kilede erinevatel külgedel erinevad omadused.
Rakumembraan (tsütolemma, plasmalemma) on kolmekihiline lipoproteiini (rasvvalk) membraan, mis eraldab iga raku naaberrakkudest ja keskkonnast ning teostab kontrollitud vahetust rakkude ja keskkonna vahel.
Selles määratluses ei ole määrav mitte see, et rakumembraan eraldaks ühe raku teisest, vaid see, et see tagab selle koostoime teiste rakkude ja keskkonnaga. Membraan on väga aktiivne, pidevalt töötav raku struktuur, millele on looduse poolt määratud palju funktsioone. Meie artiklist saate teada kõike rakumembraani koostise, struktuuri, omaduste ja funktsioonide kohta, samuti rakumembraanide talitlushäiretest tulenevate ohtude kohta inimeste tervisele.
Rakumembraanide uurimise ajalugu
1925. aastal suutsid kaks Saksa teadlast Gorter ja Grendel viia läbi kompleksse eksperimendi inimese punaste vereliblede ehk erütrotsüütidega. Osmootse šoki abil said teadlased nn "varjud" - punaste vereliblede tühjad kestad, seejärel panid need ühte hunnikusse ja mõõtsid pindala. Järgmine samm oli lipiidide hulga arvutamine rakumembraanis. Atsetooni abil eraldasid teadlased "varjudest" lipiidid ja tegid kindlaks, et neist piisab kahekordse pideva kihi jaoks.
Katse käigus tehti aga kaks jämedat viga:
Atsetooni kasutamine ei võimalda kõiki lipiide membraanidest eraldada;
"Varjude" pindala arvutati kuivkaalu järgi, mis on samuti vale.
Kuna esimene viga andis arvutustes miinuse, teine aga plussi, osutus üldtulemus üllatavalt täpseks ning Saksa teadlased tõid teadusmaailma kõige olulisema avastuse - rakumembraani lipiidide kaksikkihi.
1935. aastal jõudis teine teadlaste paar Danielly ja Dawson pärast pikki katseid bilipiidkiledega järeldusele, et rakumembraanides leidub valke. Ei olnud muud võimalust seletada, miks neil kiledel on nii suur pindpinevus. Teadlased on avalikkuse ette toonud võileivalaadse rakumembraani skemaatilise mudeli, kus saiaviilude rolli täidavad homogeensed lipiid-valgukihid ning nende vahel on õli asemel tühjus.
1950. aastal leidis esimese elektronmikroskoobi abil Danielly-Dawsoni teooria osaliselt kinnitust – rakumembraani mikrograafidel olid selgelt näha kaks lipiidi- ja valgupeadest koosnevat kihti ning nende vahel oli läbipaistev ruum, mis oli täidetud ainult lipiidide sabadega. ja valgud.
1960. aastal töötas Ameerika mikrobioloog J. Robertson nendest andmetest juhindudes välja teooria rakumembraanide kolmekihilise struktuuri kohta, mida peeti pikka aega ainsaks tõeseks. Teaduse arenedes tekkis aga üha enam kahtlusi nende kihtide homogeensuses. Termodünaamika seisukohalt on selline struktuur äärmiselt ebasoodne - rakkudel oleks väga raske transportida aineid kogu “võileiva” kaudu sisse ja välja. Lisaks on tõestatud, et erinevate kudede rakumembraanid on erineva paksuse ja kinnitusviisiga, mis on tingitud elundite erinevatest funktsioonidest.
1972. aastal leidsid mikrobioloogid S.D. Laulja ja G.L. Nicholson suutis kõik Robertsoni teooria ebakõlad selgitada rakumembraani uue vedeliku-mosaiikmudeli abil. Teadlased on leidnud, et membraan on heterogeenne, asümmeetriline, vedelikuga täidetud ja selle rakud on pidevas liikumises. Ja selle moodustavad valgud on erineva struktuuri ja eesmärgiga, lisaks paiknevad nad membraani bilipiidkihi suhtes erinevalt.
Rakumembraanid sisaldavad kolme tüüpi valke:
Perifeerne - kinnitatud kile pinnale;
poolintegraalne- tungida osaliselt läbi bilipiidkihi;
Integraalne - tungib täielikult läbi membraani.
Perifeersed valgud on elektrostaatilise interaktsiooni kaudu seotud membraani lipiidide peadega ja nad ei moodusta kunagi pidevat kihti, nagu varem arvati. Ja poolintegraalsed ja integraalsed valgud transpordivad rakku hapnikku ja toitaineid, samuti eemaldavad lagunemise. selle tooteid ja palju muud mitmete oluliste funktsioonide jaoks, millest saate hiljem teada.

Rakumembraan täidab järgmisi funktsioone:
Barjäär – membraani läbilaskvus erinevat tüüpi molekulidele ei ole sama.Rakumembraanist möödasõiduks peab molekul olema kindla suurusega, keemilised omadused ja elektrilaeng. Kahjulikud või sobimatud molekulid ei saa rakumembraani barjäärifunktsiooni tõttu lihtsalt rakku siseneda. Näiteks kaitseb membraan peroksiidreaktsiooni abil tsütoplasmat talle ohtlike peroksiidide eest;
Transport – membraani läbib passiivne, aktiivne, reguleeritud ja selektiivne vahetus. Passiivne ainevahetus sobib väga väikestest molekulidest koosnevatele rasvlahustuvatele ainetele ja gaasidele. Sellised ained tungivad difusiooni teel vabalt ilma energiakuluta rakku ja sealt välja. Vajadusel aktiveeritakse rakumembraani aktiivne transpordifunktsioon, kuid raskesti transporditavaid aineid on vaja rakku sisse või sealt välja viia. Näiteks need, kellel on suur molekulaarne suurus või mis ei suuda hüdrofoobsuse tõttu bilipiidkihti ületada. Seejärel hakkavad tööle valgupumbad, sealhulgas ATPaas, mis vastutab kaaliumiioonide imendumise eest rakku ja naatriumioonide väljutamise eest. Reguleeritud transport on oluline sekretsiooni ja fermentatsiooni funktsioonide jaoks, näiteks kui rakud toodavad ja eritavad hormoone või maomahla. Kõik need ained lahkuvad rakkudest spetsiaalsete kanalite kaudu ja etteantud mahus. Ja selektiivne transpordifunktsioon on seotud väga integreeritud valkudega, mis tungivad läbi membraani ja toimivad kanalina rangelt määratletud tüüpi molekulide sisenemiseks ja väljumiseks;
Maatriks – rakumembraan määrab ja fikseerib organellide paiknemise üksteise suhtes (tuum, mitokondrid, kloroplastid) ning reguleerib nendevahelist vastasmõju;
Mehaaniline - tagab ühe raku piiramise teisest ja samal ajal rakkude õige ühendamise homogeenseks koeks ja elundite vastupidavuse deformatsioonile;
Kaitsev - nii taimedes kui ka loomades on rakumembraan kaitseraami ehitamise aluseks. Näiteks on kõva puit, tihe koor, torkivad okkad. Loomamaailmas on ka palju näiteid rakumembraanide kaitsefunktsioonist – kilpkonna kest, kitiinkarp, kabjad ja sarved;
Energia - fotosünteesi ja rakuhingamise protsessid oleksid võimatud ilma rakumembraani valkude osaluseta, kuna rakud vahetavad energiat just valgukanalite abil;
Retseptor – rakumembraani põimitud valkudel võib olla veel üks oluline funktsioon. Need toimivad retseptoritena, mille kaudu rakk saab signaali hormoonidelt ja neurotransmitteritelt. Ja see on omakorda vajalik närviimpulsside juhtimiseks ja hormonaalsete protsesside normaalseks kulgemiseks;
Ensümaatiline - veel üks oluline funktsioon, mis on omane mõnedele rakumembraanide valkudele. Näiteks sooleepiteelis sünteesitakse selliste valkude abil seedeensüüme;
Biopotentsiaal- kaaliumiioonide kontsentratsioon rakus on palju suurem kui väljaspool ja naatriumiioonide kontsentratsioon, vastupidi, on suurem väljaspool kui sees. See seletab potentsiaalset erinevust: raku sees on laeng negatiivne, väljastpoolt positiivne, mis aitab kaasa ainete liikumisele rakku ja sealt välja ükskõik millises kolmest ainevahetustüübist – fagotsütoos, pinotsütoos ja eksotsütoos;
Märgistus - rakumembraanide pinnal on nn "märgised" - glükoproteiinidest koosnevad antigeenid (valgud, mille külge on kinnitatud hargnenud oligosahhariidsed kõrvalahelad). Kuna külgahelatel võib olla väga palju erinevaid konfiguratsioone, saab iga rakutüüp oma ainulaadse märgise, mis võimaldab teistel keharakkudel neid "nägemise järgi" ära tunda ja neile õigesti reageerida. Seetõttu tunnevad näiteks inimese immuunrakud, makrofaagid, kehasse sattunud välismaalase (infektsioon, viirus) kergesti ära ja püüavad seda hävitada. Sama juhtub haigete, muteerunud ja vanade rakkudega – nende rakumembraanil muutub silt ja keha vabaneb neist.
Rakuvahetus toimub läbi membraanide ja seda saab läbi viia kolme peamise reaktsioonitüübi kaudu:
Fagotsütoos on rakuline protsess, mille käigus membraani sisseehitatud fagotsüütrakud püüavad kinni ja seedivad toitainete tahkeid osakesi. Inimkehas viivad fagotsütoosi läbi kahte tüüpi rakkude membraanid: granulotsüüdid (granuleeritud leukotsüüdid) ja makrofaagid (immuunsuse tapjarakud);
Pinotsütoos on sellega kokkupuutuvate vedelate molekulide hõivamise protsess rakumembraani pinna kaudu. Pinotsütoosi tüübi järgi toitumiseks kasvatab rakk oma membraanile õhukesed kohevad väljakasvud antennide kujul, mis ümbritsevad justkui vedelikutilka, ja tekib mull. Esiteks ulatub see vesiikul membraani pinnast kõrgemale ja seejärel "alla neelatakse" - see peidab end raku sees ja selle seinad ühinevad rakumembraani sisepinnaga. Pinotsütoos esineb peaaegu kõigis elusrakkudes;
Eksotsütoos on pöördprotsess, mille käigus tekivad raku sees vesiikulid sekretoorse funktsionaalse vedelikuga (ensüüm, hormoon), mis tuleb kuidagi rakust keskkonda eemaldada. Selleks sulandub mull esmalt rakumembraani sisepinnaga, seejärel pundub väljapoole, lõhkeb, ajab sisu välja ja sulandub uuesti membraani pinnaga, seekord väljastpoolt. Eksotsütoos toimub näiteks sooleepiteeli ja neerupealiste koore rakkudes.
Kas leidsite tekstist vea? Valige see ja veel paar sõna, vajutage Ctrl + Enter

Rakumembraanid sisaldavad kolme lipiidide klassi:
fosfolipiidid;
glükolipiidid;
Kolesterool.
Fosfolipiidid (rasvade ja fosfori kombinatsioon) ja glükolipiidid (rasvade ja süsivesikute kombinatsioon) koosnevad omakorda hüdrofiilsest peast, millest ulatub välja kaks pikka hüdrofoobset saba. Kuid kolesterool hõivab mõnikord nende kahe saba vahelise ruumi ega lase neil painduda, mis muudab mõne raku membraanid jäigaks. Lisaks muudavad kolesterooli molekulid sujuvamaks rakumembraanide struktuuri ja takistavad polaarsete molekulide üleminekut ühest rakust teise.
Kuid kõige olulisem komponent, nagu võib näha eelmisest rakumembraanide funktsioone käsitlevast jaotisest, on valgud. Nende koostis, otstarve ja asukoht on väga mitmekesised, kuid on midagi ühist, mis neid kõiki ühendab: rõngakujulised lipiidid paiknevad alati rakumembraanide valkude ümber. Need on spetsiaalsed rasvad, mis on selgelt struktureeritud, stabiilsed, nende koostises on rohkem küllastunud rasvhappeid ja mis vabanevad membraanidest koos "sponsoreeritud" valkudega. See on omamoodi isiklik kaitsekesta valkude jaoks, ilma milleta need lihtsalt ei töötaks.
Rakumembraani struktuur on kolmekihiline. Keskel on suhteliselt homogeenne vedel bilipiidkiht ja valgud katavad seda mõlemalt poolt omamoodi mosaiigiga, tungides osaliselt paksusesse. See tähendab, et oleks vale arvata, et rakumembraanide välimised valgukihid on pidevad. Valgud on lisaks nende keerukatele funktsioonidele vajalikud membraanis selleks, et rakkudesse siseneda ja sealt välja transportida need ained, mis ei suuda rasvakihti tungida. Näiteks kaaliumi- ja naatriumioonid. Nende jaoks on ette nähtud spetsiaalsed valgustruktuurid - ioonkanalid, mida käsitleme üksikasjalikumalt allpool.
Kui vaadata rakumembraani läbi mikroskoobi, on näha kõige väiksematest sfäärilistest molekulidest moodustunud lipiidide kihti, mida mööda hõljuvad sarnaselt merega suured erineva kujuga valgurakud. Täpselt samad membraanid jagavad iga raku siseruumi osadeks, milles tuum, kloroplastid ja mitokondrid mugavalt paiknevad. Kui raku sees ei oleks eraldi “ruume”, kleepuksid organellid kokku ega suudaks oma funktsioone õigesti täita.
Rakk on membraanidega struktureeritud ja piiritletud organellide kogum, mis osaleb energia-, ainevahetus-, informatsiooni- ja paljunemisprotsesside kompleksis, mis tagavad organismi elutegevuse.
Nagu sellest määratlusest näha, on membraan iga raku kõige olulisem funktsionaalne komponent. Selle tähtsus on sama suur kui tuuma, mitokondrite ja teiste rakuorganellide oma. Ja membraani ainulaadsed omadused tulenevad selle struktuurist: see koosneb kahest erilisel viisil kokkukleepunud kilest. Fosfolipiidide molekulid membraanis paiknevad hüdrofiilsete peadega väljapoole ja hüdrofoobsete sabadega sissepoole. Seetõttu on kile üks pool veest märjaks, teine aga mitte. Niisiis on need kiled üksteisega ühendatud mittemärguvate külgedega sissepoole, moodustades bilipiidkihi, mida ümbritsevad valgumolekulid. See on rakumembraani "sandwich" struktuur.
Rakumembraanide ioonikanalid
Vaatleme üksikasjalikumalt ioonkanalite tööpõhimõtet. Milleks neid vaja on? Fakt on see, et läbi lipiidmembraani võivad vabalt tungida ainult rasvlahustuvad ained - need on gaasid, alkoholid ja rasvad ise. Nii näiteks toimub punastes verelibledes pidev hapniku ja süsihappegaasi vahetus ning selleks ei pea meie keha kasutama mingeid täiendavaid nippe. Mis saab aga siis, kui on vaja transportida vesilahuseid, nagu naatriumi- ja kaaliumisoolad, läbi rakumembraani?
Sellistele ainetele oleks bilipiidkihis võimatu teed sillutada, kuna augud tõmbuksid kohe kokku ja kleepuksid kokku tagasi, selline on iga rasvkoe struktuur. Kuid loodus, nagu alati, leidis olukorrast väljapääsu ja lõi spetsiaalsed valgu transpordistruktuurid.
Juhtivaid valke on kahte tüüpi:
Transporterid on poolintegreeritud valgupumbad;
Kanaloformerid on lahutamatud valgud.
Esimest tüüpi valgud on osaliselt sukeldatud rakumembraani bilipiidkihti ja vaatavad oma peaga välja ning soovitud aine juuresolekul hakkavad nad käituma nagu pump: tõmbavad molekuli ligi ja imevad selle endasse. kamber. Ja teist tüüpi, integraalsed valgud on pikliku kujuga ja asuvad rakumembraani bilipiidkihiga risti, tungides selle läbi ja läbi. Nende kaudu, nagu ka tunnelite kaudu, liiguvad rakku sisse ja sealt välja ained, mis ei suuda rasvast läbi minna. Ioonikanalite kaudu tungivad kaaliumiioonid rakku ja kogunevad sinna, naatriumioonid aga vastupidi. Elektrilistes potentsiaalides on erinevus, mis on nii vajalik meie keha kõigi rakkude nõuetekohaseks toimimiseks.
Olulisemad järeldused rakumembraanide ehituse ja funktsioonide kohta

Teooria tundub alati huvitav ja paljutõotav, kui seda saab praktikas kasulikult rakendada. Inimkeha rakumembraanide struktuuri ja funktsioonide avastamine võimaldas teadlastel teha tõelise läbimurde teaduses üldiselt ja eriti meditsiinis. Pole juhus, et oleme ioonkanalitel nii üksikasjalikult peatunud, sest just siin peitub vastus meie aja ühele kõige olulisemale küsimusele: miks haigestuvad inimesed üha sagedamini onkoloogiasse?
Vähk nõuab igal aastal maailmas umbes 17 miljonit inimelu ja on kõigi surmade põhjuste hulgas neljas. WHO andmetel kasvab vähki haigestumine pidevalt ning 2020. aasta lõpuks võib see ulatuda 25 miljonini aastas.
Mis seletab tõelist vähiepideemiat ja kuidas on sellega pistmist rakumembraanide funktsioon? Te ütlete: põhjuseks on halvad keskkonnatingimused, alatoitumus, halvad harjumused ja raske pärilikkus. Ja muidugi on teil õigus, aga kui probleemist lähemalt rääkida, siis põhjuseks on inimkeha hapestumine. Eespool loetletud negatiivsed tegurid põhjustavad rakumembraanide häireid, pärsivad hingamist ja toitumist.
Seal, kus peaks olema pluss, tekib miinus ja rakk ei saa normaalselt toimida. Kuid vähirakud ei vaja ei hapnikku ega leeliselist keskkonda - nad on võimelised kasutama anaeroobset tüüpi toitumist. Seetõttu muteeruvad terved rakud hapnikuvaeguse ja skaalavälise pH-taseme tingimustes, soovides keskkonnaga kohaneda ja muutuvad vähirakkudeks. Nii haigestub inimene vähki. Selle vältimiseks peate lihtsalt iga päev jooma piisavalt puhast vett ja loobuma kantserogeenidest toidus. Kuid reeglina on inimesed kahjulikest toodetest ja kvaliteetse vee vajadusest teadlikud ega tee midagi - nad loodavad, et hädad lähevad neist mööda.
Teades erinevate rakkude rakumembraanide struktuuri ja funktsioonide omadusi, saavad arstid seda teavet kasutada, et pakkuda kehale sihipärast ja sihipärast ravitoimet. Paljud kaasaegsed ravimid, sattudes meie kehasse, otsivad õiget "sihtmärki", milleks võivad olla rakumembraanide ioonkanalid, ensüümid, retseptorid ja biomarkerid. See ravimeetod võimaldab saavutada paremaid tulemusi minimaalsete kõrvalmõjudega.
Viimase põlvkonna antibiootikumid ei tapa verre sattudes kõiki rakke järjest, vaid otsivad täpselt üles patogeeni rakud, keskendudes selle rakumembraanides olevatele markeritele. Uusimad migreenivastased ravimid, triptaanid, ahendavad ainult aju põletikulisi veresooni, avaldamata peaaegu mingit mõju südamele ja perifeersele vereringesüsteemile. Ja nad tunnevad vajalikud veresooned ära täpselt oma rakumembraanide valkude järgi. Selliseid näiteid on palju, seega võime kindlalt väita, et teadmised rakumembraanide ehituse ja funktsioonide kohta on kaasaegse arstiteaduse arengu aluseks ning päästavad igal aastal miljoneid elusid.
Artikli autor: meditsiiniteaduste doktor, terapeut Mochalov Pavel Aleksandrovich
Kamber- see pole mitte ainult vedelik, ensüümid ja muud ained, vaid ka kõrgelt organiseeritud struktuurid, mida nimetatakse intratsellulaarseteks organellideks. Raku organellid pole vähem tähtsad kui selle keemilised komponendid. Seega, kui puuduvad organellid nagu mitokondrid, väheneb toitainetest eraldatud energiavaru koheselt 95%.
Enamik raku organelle on kaetud membraanid koosneb peamiselt lipiididest ja valkudest. Seal on rakkude membraanid, endoplasmaatiline retikulum, mitokondrid, lüsosoomid, Golgi aparaat.
Lipiidid on vees lahustumatud, mistõttu nad loovad rakus barjääri, mis takistab vee ja vees lahustuvate ainete liikumist ühest sektsioonist teise. Valgumolekulid aga muudavad membraani läbi spetsiaalsete struktuuride, mida nimetatakse poorideks, läbilaskvaks erinevatele ainetele. Paljud teised membraanivalgud on ensüümid, mis katalüüsivad arvukalt keemilisi reaktsioone, mida arutatakse järgmistes peatükkides.
Raku (või plasma) membraan on õhuke, painduv ja elastne struktuur paksusega vaid 7,5-10 nm. See koosneb peamiselt valkudest ja lipiididest. Selle komponentide ligikaudne suhe on järgmine: valgud - 55%, fosfolipiidid - 25%, kolesterool - 13%, muud lipiidid - 4%, süsivesikud - 3%.
rakumembraani lipiidkiht takistab vee läbitungimist. Membraani aluseks on lipiidne kaksikkiht - õhuke lipiidkile, mis koosneb kahest monokihist ja katab raku täielikult. Kogu membraanis on valgud suurte gloobulite kujul.
lipiidide kaksikkiht koosneb peamiselt fosfolipiidi molekulidest. Sellise molekuli üks ots on hüdrofiilne, s.t. vees lahustuv (sellel asub fosfaatrühm), teine on hüdrofoobne, s.t. lahustub ainult rasvades (sisaldab rasvhapet).
Tulenevalt asjaolust, et molekuli hüdrofoobne osa fosfolipiid tõrjub vett, kuid neid tõmbavad samade molekulide sarnased osad, on fosfolipiididel loomulik omadus kinnituda üksteise külge membraani paksuses, nagu on näidatud joonisel fig. 2-3. Fosfaatrühmaga hüdrofiilne osa moodustab kaks membraanipinda: välimise, mis on kontaktis rakuvälise vedelikuga, ja sisemise, mis on kontaktis rakusisese vedelikuga.
Keskmine lipiidikiht ioonide ning glükoosi ja uurea vesilahuste suhtes läbimatu. Rasvlahustuvad ained, sealhulgas hapnik, süsinikdioksiid, alkohol, vastupidi, tungivad kergesti sellesse membraani piirkonda.
molekulid Kolesterool, mis on membraani osa, on samuti loomulikult lipiidid, kuna nende steroidrühmal on kõrge lahustuvus rasvades. Need molekulid näivad olevat lahustunud lipiidide kaksikkihis. Nende põhieesmärk on reguleerida kehavedelike vees lahustuvate komponentide membraanide läbilaskvust (või mitteläbilaskvust). Lisaks on kolesterool membraani viskoossuse peamine regulaator.
Rakumembraani valgud. Joonisel on lipiidide kaksikkihis nähtavad kerakujulised osakesed – need on membraanivalgud, millest enamus on glükoproteiinid. Membraanvalke on kahte tüüpi: (1) integraalsed, mis tungivad läbi membraani; (2) perifeersed, mis ulatuvad välja ainult ühe pinna kohal, teisele jõudmata.
Paljud integraalsed valgud moodustavad kanaleid (või poore), mille kaudu vesi ja vees lahustuvad ained, eriti ioonid, võivad difundeeruda rakusise- ja ekstratsellulaarsesse vedelikku. Kanalite selektiivsuse tõttu hajuvad mõned ained paremini kui teised.
Muud integraalsed valgud toimivad kandevalkudena, juhtides ainete transporti, mille lipiidide kaksikkiht on läbitungimatu. Mõnikord toimivad kandjavalgud difusioonile vastupidises suunas, sellist transporti nimetatakse aktiivseks. Mõned integraalsed valgud on ensüümid.
Integraalsed membraanivalgud võivad toimida ka veeslahustuvate ainete, sealhulgas peptiidhormoonide retseptoritena, kuna membraan on neile mitteläbilaskev. Retseptorvalgu interaktsioon teatud ligandiga põhjustab valgu molekulis konformatsioonilisi muutusi, mis omakorda stimuleerivad valgu molekuli rakusisese segmendi ensümaatilist aktiivsust või signaali ülekandmist retseptorist rakku, kasutades teist sõnumitoojat. Seega kaasavad rakumembraani sisseehitatud integraalsed valgud selle väliskeskkonna info rakku edastamise protsessi.
Perifeersete membraanide valkude molekulid seostatakse sageli integraalsete valkudega. Enamik perifeersetest valkudest on ensüümid või täidavad dispetšeri rolli ainete transportimisel läbi membraanipooride.
_Teemad _
4.1. Membraanide üldised omadused. Membraanide struktuur ja koostis
4.2. Ainete transport läbi membraanide
4.3. Transmembraanne signaalimine _
Õppeeesmärgid Et oleks võimalik:
1. Tõlgendada membraanide rolli ainevahetuse reguleerimisel, ainete transportimisel rakku ja metaboliitide eemaldamisel.
2. Selgitage hormoonide ja teiste signaalmolekulide molekulaarseid toimemehhanisme sihtorganitel.
Tea:
1. Bioloogiliste membraanide ehitus ja roll ainevahetuses ja energias.
2. Peamised viisid ainete ülekandmiseks läbi membraanide.
3. Hormoonide, vahendajate, tsütokiinide, eikosanoidide transmembraanse signaaliülekande põhikomponendid ja etapid.
TEEMA 4.1. MEMBRAANI ÜLDOMADUSED.
MEMBRAANIDE STRUKTUUR JA KOOSTIS
Kõik rakud ja intratsellulaarsed organellid on ümbritsetud membraanidega, millel on oluline roll nende struktuurilises organiseerimises ja toimimises. Kõigi membraanide ehituspõhimõtted on samad. Plasmamembraanil, aga ka endoplasmaatilise retikulumi membraanidel, Golgi aparaadil, mitokondritel ja tuumal on aga olulised struktuursed tunnused, need on ainulaadsed oma koostise ja funktsioonide olemuse poolest.
Membraan:
• eraldada rakud keskkonnast ja jagada see sektsioonideks (compartments);
• reguleerida ainete transporti rakkudesse ja organellidesse ja vastupidi;
• pakkuda rakkudevaheliste kontaktide spetsiifilisust;
• tajuda väliskeskkonna signaale.
Membraansüsteemide, sealhulgas retseptorite, ensüümide, transpordisüsteemide koordineeritud toimimine aitab säilitada raku homöostaasi ja reageerida kiiresti väliskeskkonna seisundi muutustele, reguleerides rakkudesisest ainevahetust.
Bioloogilised membraanid koosnevad lipiididest ja valkudest, mis on omavahel seotud mittekovalentne interaktsioonid. Membraani alus on kahekordne lipiidikiht mis sisaldab valgumolekule (joon. 4.1). Lipiidide kaksikkiht koosneb kahest reast amfifiilsed molekulid, mille hüdrofoobsed "sabad" on peidetud sees, ja hüdrofiilsed rühmad - polaarsed "pead" on pööratud väljapoole ja puutuvad kokku vesikeskkonnaga.
1. Membraani lipiidid. Membraanlipiidid sisaldavad nii küllastunud kui ka küllastumata rasvhappeid. Küllastumata rasvhapped on kaks korda tavalisemad kui küllastunud rasvhapped, mis määrab voolavus membraanid ja membraanivalkude konformatsiooniline labiilsus.
Membraanides on kolm peamist tüüpi lipiide – fosfolipiidid, glükolipiidid ja kolesterool (joonis 4.2 – 4.4). Kõige sagedamini leitud Glütserofosfolipiidid on fosfatiidhappe derivaadid.
Riis. 4.1. Plasmamembraani ristlõige
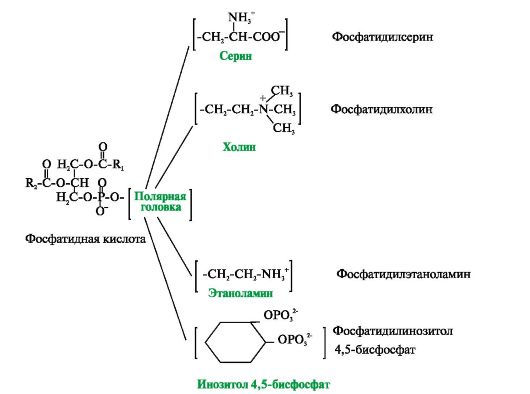 Riis. 4.2. Glütserofosfolipiidid.
Riis. 4.2. Glütserofosfolipiidid.
Fosfatiidhape on diatsüülglütseroolfosfaat. R 1 , R 2 - rasvhapperadikaalid (hüdrofoobsed "sabad"). Polüküllastumata rasvhappejääk on seotud glütserooli teise süsinikuaatomiga. Polaarne "pea" on fosforhappe jääk ja selle külge kinnitatud hüdrofiilne seriini, koliini, etanoolamiini või inositooli rühm
Samuti on lipiidid - derivaadid aminoalkohol sfingosiin.
Aminoalkohol sfingosiin atsüülimisel, st. sidudes rasvhappe NH 2 rühmaga, muutub keramiidiks. Keramiide eristab nende rasvhappejääk. Keramiidi OH-rühmaga võib seostada erinevaid polaarseid rühmi. Sõltuvalt polaarse "pea" struktuurist jagatakse need derivaadid kahte rühma - fosfolipiidid ja glükolipiidid. Sfingofosfolipiidide (sfingomüeliinide) polaarse rühma struktuur on sarnane glütserofosfolipiididega. Närvikiudude müeliinkestades leidub palju sfingomüeliine. Glükolipiidid on keramiidi süsivesikute derivaadid. Sõltuvalt süsivesikute komponendi struktuurist eristatakse tserebrosiide ja gangliosiide.
kolesterooli leidub kõigi loomarakkude membraanides, see jäigastab membraane ja vähendab neid voolavus(voolavus). Kolesterooli molekul asub membraani hüdrofoobses tsoonis paralleelselt fosfo- ja glükolipiidimolekulide hüdrofoobsete "sabadega". Kolesterooli hüdroksüülrühm, samuti fosfo- ja glükolipiidide hüdrofiilsed "pead",
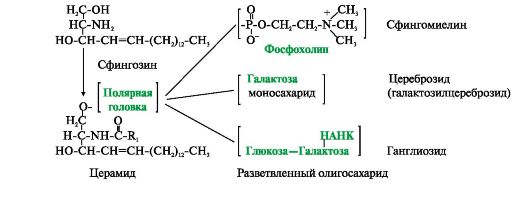 Riis. 4.3. Aminoalkoholi sfingosiini derivaadid.
Riis. 4.3. Aminoalkoholi sfingosiini derivaadid.
Keramiid – atsüülitud sfingosiin (R 1 – rasvhapperadikaal). Fosfolipiidide hulka kuuluvad sfingomüeliinid, milles polaarne rühm koosneb fosforhappejäägist ja koliinist, etanoolamiinist või seriinist. Glükolipiidide hüdrofiilne rühm (polaarne "pea") on süsivesikute jääk. Tserebrosiidid sisaldavad lineaarset mono- või oligosahhariidi jääki. Gangliosiidide koostis sisaldab hargnenud oligosahhariidi, mille üks monomeerseid ühikuid on NANK - N-atsetüülneuramiinhape
näoga veefaasi poole. Kolesterooli ja teiste lipiidide molaarsuhe membraanides on 0,3-0,9. Sellel väärtusel on tsütoplasmaatilise membraani jaoks kõrgeim väärtus.
Kolesterooli sisalduse suurenemine membraanides vähendab rasvhappeahelate liikuvust, mis mõjutab membraanivalkude konformatsioonilist labiilsust ja vähendab nende tekkevõimalust. külgmine difusioon. Membraanide voolavuse suurenemisega, mis on põhjustatud lipofiilsete ainete toimest neile või lipiidide peroksüdatsioonist, suureneb kolesterooli osakaal membraanides.
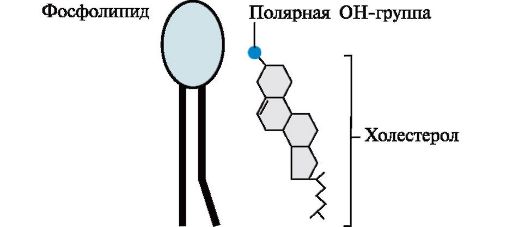 Riis. 4.4. Asukoht fosfolipiidide ja kolesterooli membraanis.
Riis. 4.4. Asukoht fosfolipiidide ja kolesterooli membraanis.
Kolesterooli molekul koosneb jäigast hüdrofoobsest tuumast ja painduvast süsivesinikahelast. Polaarne "pea" on OH-rühm kolesterooli molekuli 3. süsinikuaatomi juures. Võrdluseks on joonisel membraani fosfolipiidi skemaatiline esitus. Nende molekulide polaarpea on palju suurem ja sellel on laeng
Membraanide lipiidide koostis on erinev, ühe või teise lipiidi sisalduse määravad ilmselt mitmesugused funktsioonid, mida need molekulid membraanides täidavad.
Membraani lipiidide peamised funktsioonid on järgmised:
• moodustada lipiidne kaksikkiht – membraanide struktuurne alus;
• tagada membraanivalkude toimimiseks vajalik keskkond;
• osaleda ensüümi aktiivsuse reguleerimises;
• toimida pinnavalkude "ankruna";
• osaleda hormonaalsete signaalide edastamises.
Muutused lipiidide kaksikkihi struktuuris võivad põhjustada membraani funktsioonide häireid.
2. Membraanvalgud. Membraanvalgud erinevad oma asukoha poolest membraanis (joon. 4.5). Lipiidide kaksikkihi hüdrofoobse piirkonnaga kontaktis olevad membraanvalgud peavad olema amfifiilsed, s.t. neil on mittepolaarne domeen. Amfifiilsus saavutatakse tänu sellele, et:
• lipiidide kaksikkihiga kontaktis olevad aminohappejäägid on enamasti mittepolaarsed;
• Paljud membraanivalgud on kovalentselt seotud rasvhappejääkidega (atsüülitud).
Valgu külge kinnitatud rasvhapete atsüüljäägid tagavad selle "ankurdumise" membraanis ja külgmise difusiooni võimaluse. Lisaks läbivad membraanivalgud translatsioonijärgseid modifikatsioone, nagu glükosüülimine ja fosforüülimine. Integraalsete valkude välispinna glükosüülimine kaitseb neid rakkudevahelise ruumi proteaaside kahjustuste eest.
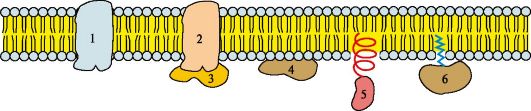 Riis. 4.5. Membraanvalgud:
Riis. 4.5. Membraanvalgud:
1, 2 - integraalsed (transmembraansed) valgud; 3, 4, 5, 6 - pinnavalgud. Integraalsetes valkudes on osa polüpeptiidahelast põimitud lipiidikihti. Need valgu osad, mis interakteeruvad rasvhapete süsivesinike ahelatega, sisaldavad valdavalt mittepolaarseid aminohappeid. Polaarsete "peade" piirkonnas asuvad valgu piirkonnad on rikastatud hüdrofiilsete aminohappejääkidega. Pinnavalgud kinnituvad membraanile erineval viisil: 3 - seotud integraalsete valkudega; 4 - kinnitatud lipiidikihi polaarsete "peade" külge; 5 - "ankurdatud" membraanis lühikese hüdrofoobse terminaalse domeeniga; 6 - "ankurdatud" membraani, kasutades kovalentselt seotud atsüüljääki
Sama membraani välimine ja sisemine kiht erinevad lipiidide ja valkude koostise poolest. Seda funktsiooni membraanide struktuuris nimetatakse transmembraanne asümmeetria.
Membraanvalgud võivad olla seotud:
• ainete selektiivne transport rakku ja sealt välja;
• hormonaalsete signaalide edastamine;
• endotsütoosi ja eksotsütoosiga seotud "piiratud süvendite" moodustumine;
• immunoloogilised reaktsioonid;
• ensüümidena ainete muundumisel;
• kudede ja elundite moodustumist tagavate rakkudevaheliste kontaktide organiseerimine.
TEEMA 4.2. AINETE TRANSPORT LÄBI MEMBRAANI
Membraanide üks põhifunktsioone on ainete rakku sisse ja välja liikumise reguleerimine, rakule vajalike ainete kinnipidamine ja ebavajalike eemaldamine. Ioonide, orgaaniliste molekulide transport läbi membraanide võib toimuda mööda kontsentratsioonigradienti - passiivne transport ja kontsentratsiooni gradiendi vastu - aktiivne transport.
1. Passiivne transport saab teostada järgmistel viisidel (joonis 4.6, 4.7):
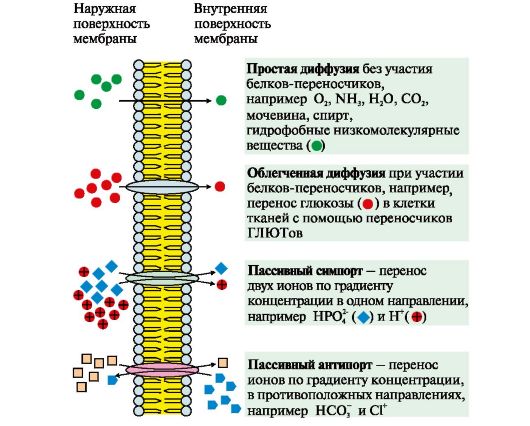 Riis. 4.6. Ainete ülekandumise mehhanismid läbi membraanide piki kontsentratsioonigradienti
Riis. 4.6. Ainete ülekandumise mehhanismid läbi membraanide piki kontsentratsioonigradienti
Passiivne transport on ioonide difusioon läbi valgukanalite, näiteks H+, Ca 2+, N+, K+ difusioon. Enamiku kanalite toimimist reguleerivad spetsiifilised ligandid või transmembraanse potentsiaali muutused.
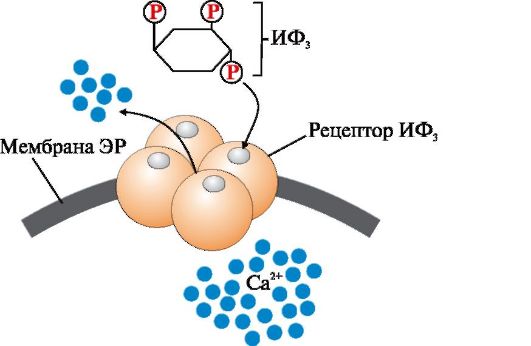 Riis. 4.7. Endoplasmaatilise retikulumi membraani Ca2+ kanal, mida reguleerib inositool-1,4,5-trifosfaat (IF 3).
Riis. 4.7. Endoplasmaatilise retikulumi membraani Ca2+ kanal, mida reguleerib inositool-1,4,5-trifosfaat (IF 3).
IP 3 (inositool-1,4,5-trifosfaat) moodustub membraani lipiidi PIF 2 (fosfatidüülinositool-4,5-bisfosfaat) hüdrolüüsil ensüümi fosfolipaas C toimel. IP 3 seondub membraani spetsiifiliste keskustega. endoplasmaatilise retikulumi membraani Ca 2 + kanali protomeerid. Valgu konformatsioon muutub ja kanal avaneb - Ca 2 + siseneb raku tsütosooli piki kontsentratsioonigradienti
2. Aktiivne transport. esmane aktiivne transport toimub kontsentratsioonigradienti vastu ATP energia kulutamisel transpordi-ATPaaside osalusel, näiteks Na +, K + -ATPaas, H + -ATPaas, Ca 2 + -ATPaas (joon. 4.8). H + -ATPaasid toimivad prootonpumpadena, mis loovad raku lüsosoomides happelise keskkonna. Tsütoplasmaatilise membraani ja endoplasmaatilise retikulumi membraani Ca 2+ -ATPaasi abil hoitakse madalat kaltsiumi kontsentratsiooni raku tsütosoolis ning luuakse rakusisene Ca 2+ depoo mitokondrites ja endoplasmaatilises membraanis. võrkkest.
sekundaarne aktiivne transport toimub ühe transporditava aine (joonis 4.9) kontsentratsioonigradiendi tõttu, mille tekitab kõige sagedamini Na +, K + -ATPaas, mis toimib koos ATP tarbimisega.
Suurema kontsentratsiooniga aine kinnitumine kandevalgu aktiivtsentri külge muudab selle konformatsiooni ja suurendab afiinsust rakku läbiva ühendi suhtes kontsentratsioonigradiendi suhtes. Sekundaarset aktiivset transporti on kahte tüüpi: aktiivne sümptom ja antiport.
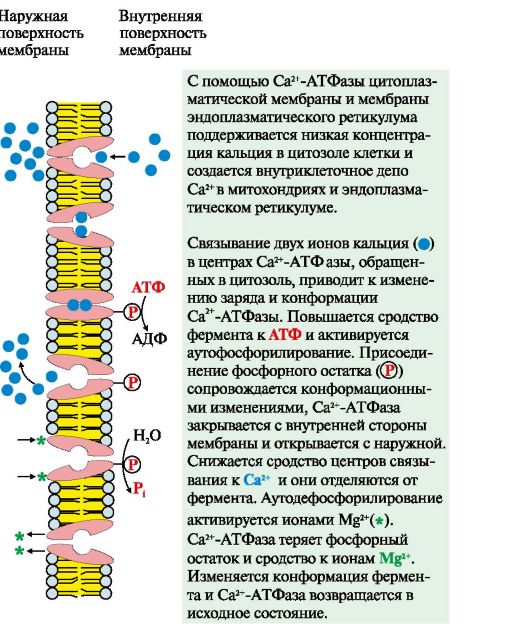 Riis. 4.8. Ca 2 + -ATPaasi toimimise mehhanism
Riis. 4.8. Ca 2 + -ATPaasi toimimise mehhanism
 Riis. 4.9. sekundaarne aktiivne transport
Riis. 4.9. sekundaarne aktiivne transport
3. Makromolekulide ja osakeste ülekanne membraanide osalusel - endotsütoos ja eksotsütoos.
Makromolekulide, näiteks valkude, nukleiinhapete, polüsahhariidide või isegi suuremate osakeste ülekandumine rakuvälisest keskkonnast rakku toimub endotsütoos. Ainete või kõrgmolekulaarsete komplekside sidumine toimub plasmamembraani teatud piirkondades, mida nimetatakse vooderdatud süvendid. Endotsütoos, mis toimub ääristatud aukudesse ehitatud retseptorite osalusel, võimaldab rakkudel absorbeerida spetsiifilisi aineid ja seda nimetatakse retseptorist sõltuv endotsütoos.
Makromolekulid, nagu peptiidhormoonid, seedeensüümid, ekstratsellulaarsed maatriksvalgud, lipoproteiinikompleksid, erituvad verre või rakkudevahelisse ruumi eksotsütoos. See transpordiviis võimaldab eemaldada rakust sekretoorsetes graanulites kogunevaid aineid. Enamikul juhtudel reguleeritakse eksotsütoosi kaltsiumiioonide kontsentratsiooni muutmisega rakkude tsütoplasmas.
TEEMA 4.3. TRANSMEMBRAANNE SIGNAALID
Membraanide oluline omadus on võime tajuda ja edastada rakusisese keskkonna signaale. Rakud tajuvad väliseid signaale, kui nad interakteeruvad sihtrakkude membraanis asuvate retseptoritega. Retseptorid, ühendades signaalmolekuli, aktiveerivad rakusiseseid infoülekandeteid, mis viib erinevate metaboolsete protsesside kiiruse muutumiseni.
1. Signaalmolekul, mis interakteerub spetsiifiliselt membraaniretseptoriga esmane sõnumitooja. Primaarsete sõnumikandjatena toimivad mitmesugused keemilised ühendid – hormoonid, neurotransmitterid, eikosanoidid, kasvufaktorid või füüsikalised tegurid, näiteks valguskvant. Primaarsete sõnumitoojate poolt aktiveeritud rakumembraani retseptorid edastavad saadud teabe valkude ja ensüümide süsteemi, mis moodustab signaali edastamise kaskaad, signaali võimenduse mitusada korda. Raku reaktsiooniaeg, mis seisneb ainevahetusprotsesside aktiveerimises või inaktiveerimises, lihaskontraktsioonis, ainete transportimises sihtrakkudest, võib olla mitu minutit.
Membraan retseptorid jagatud:
• retseptorid, mis sisaldavad subühikut, mis seob primaarset sõnumitoojat ja ioonkanalit;
• retseptorid, mis on võimelised avaldama katalüütilist aktiivsust;
• retseptorid, G-valkude abil aktiveerides sekundaarsete (rakusisese) sõnumitoojate moodustumist, mis edastavad signaali tsütosooli spetsiifilistele valkudele ja ensüümidele (joonis 4.10).
Teised sõnumitoojad on väikese molekulmassiga, difundeeruvad suure kiirusega raku tsütosoolis, muudavad vastavate valkude aktiivsust ja seejärel jagunevad kiiresti või eemaldatakse tsütosoolist.
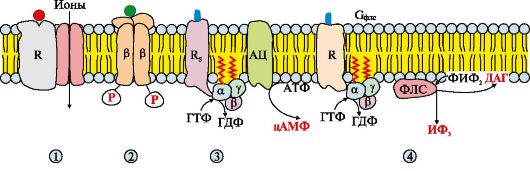 Riis. 4.10. Membraanis asuvad retseptorid.
Riis. 4.10. Membraanis asuvad retseptorid.
Membraani retseptorid võib jagada kolme rühma. Retseptorid: 1 - sisaldavad subühikut, mis seob signaalmolekuli ja ioonikanalit, näiteks atsetüülkoliini retseptorit postsünaptilisel membraanil; 2 - katalüütiline aktiivsus pärast signaalmolekuli, näiteks insuliiniretseptori lisamist; 3, 4 - signaali edastamine ensüümile adenülaattsüklaas (AC) või fosfolipaas C (PLS) membraani G-valkude, näiteks erinevat tüüpi adrenaliini, atsetüülkoliini ja muude signaalmolekulide retseptorite osalusel.
Roll sekundaarsed sõnumitoojad teostada molekule ja ioone:
• cAMP (tsükliline adenosiin-3",5"-monofosfaat);
• cGMP (tsükliline guanosiin-3",5"-monofosfaat);
• IF 3 (inositool-1,4,5-trifosfaat);
• DAG (diatsüülglütserool);
Seal on hormoonid (steroid ja kilpnääre), mis läbides lipiidide kaksikkihti, lahtrisse siseneda ja suhelda rakusisesed retseptorid. Füsioloogiliselt oluline erinevus membraani ja intratsellulaarsete retseptorite vahel on vastuse kiirus sissetulevale signaalile. Esimesel juhul on mõju kiire ja lühiajaline, teisel - aeglane, kuid kauakestev.
G-valguga seotud retseptorid
Hormoonide interaktsioon G-valguga seotud retseptoritega põhjustab inositoolfosfaadi signaaliülekandesüsteemi aktiveerumist või muutusi adenülaattsüklaasi regulatsioonisüsteemi aktiivsuses.
2. Adenülaattsüklaasi süsteem sisaldab (joonis 4.11):
- lahutamatu tsütoplasma membraani valgud:
• Rs - primaarse sõnumitooja retseptor - adenülaattsüklaasi süsteemi (ACS) aktivaator;
• R ; - primaarse sõnumikandja retseptor - ACS-i inhibiitor;
• ensüüm adenülaattsüklaas (AC).
- "ankurdatud" valgud:
• G s - GTP-d siduv valk, mis koosneb α,βγ-subühikutest, milles (α,-subühik on seotud GDP molekuliga;
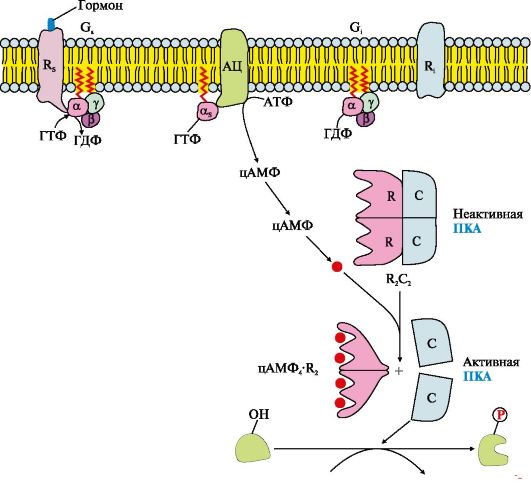
Riis. 4.11. Adenülaattsüklaasi süsteemi toimimine
• G ; - GTP-d siduv valk, mis koosneb αβγ-subühikutest, milles a; -subühik on seotud SKT molekuliga; - tsütosoolne proteiinkinaas A (PKA) ensüüm.
Adenülaattsüklaasi süsteemi primaarse messenger-signaali ülekande sündmuste jada
Retseptoril on membraani välispinnal primaarse messengeri ja G-valgu (α,βγ-GDP) seondumiskohad membraani sisepinnal. Adenülaattsüklaasi süsteemi aktivaatori, näiteks hormooni koostoime retseptoriga (Rs) viib retseptori konformatsiooni muutumiseni. Retseptori afiinsus G..-valgu suhtes suureneb. Hormoon-retseptori kompleksi sidumine GS-GDP-ga vähendab G..-valgu α,-subühiku afiinsust GDP suhtes ja suurendab afiinsust GTP suhtes. α,-subühiku aktiivses kohas asendatakse GDP GTP-ga. See põhjustab muutusi α-subühiku konformatsioonis ja selle afiinsuse vähenemist βγ-subühikute suhtes. Eraldunud subühik α,-GTP liigub membraani lipiidkihis külgsuunas ensüümi poole. adenülaattsüklaas.
α,-GTP interaktsioon adenülaattsüklaasi regulatoorse tsentriga muudab ensüümi konformatsiooni, viib selle aktiveerimiseni ja teise sõnumitooja - tsüklilise adenosiin-3,5'-monofosfaadi (cAMP) moodustumise kiiruse suurenemiseni. ATP-st. cAMP kontsentratsioon rakus suureneb. cAMP molekulid võivad pöörduvalt seonduda proteiinkinaasi A (PKA) regulatoorsete alaühikutega, mis koosneb kahest regulatoorsest (R) ja kahest katalüütilisest (C) subühikust – (R 2 C 2). Kompleks R2C2 ei oma ensümaatilist aktiivsust. cAMP-i kinnitamine regulatoorsete allüksustega põhjustab muutusi nende konformatsioonis ja komplementaarsuse kadumise C-subühikutega. Katalüütilised subühikud omandavad ensümaatilise aktiivsuse.
Aktiivne proteiinkinaas A fosforüülib ATP abil spetsiifilisi valke seriini ja treoniini jääkide juures. Valkude ja ensüümide fosforüülimine suurendab või vähendab nende aktiivsust, mistõttu muutub nende osalemise metaboolsete protsesside kiirus.
R-retseptori signaalmolekuli aktiveerimine stimuleerib Gj-valgu talitlust, mis toimub samade reeglite järgi nagu G..-valgu puhul. Kuid kui αi-GTP subühik interakteerub adenülaattsüklaasiga, väheneb ensüümi aktiivsus.
Adenülaattsüklaasi ja proteiinkinaasi A inaktiveerimine
α,-subühik kompleksis GTP-ga hakkab adenülaattsüklaasiga interakteerudes avaldama ensümaatilist (GTP-fosfataasi) aktiivsust, hüdrolüüsib GTP-d. Saadud GDP molekul jääb α-subühiku aktiivsesse keskusesse, muudab selle konformatsiooni ja vähendab selle afiinsust vahelduvvoolu suhtes. AC ja α,-GDP kompleks dissotsieerub, α,-GDP sisaldub G..-valgus. α,-GDP eraldamine adenülaattsüklaasist inaktiveerib ensüümi ja peatab cAMP sünteesi.
Fosfodiesteraas- tsütoplasmaatilise membraani "ankurdatud" ensüüm hüdrolüüsib eelnevalt moodustunud cAMP molekulid AMP-ks. cAMP kontsentratsiooni vähenemine rakus põhjustab cAMP 4 K " 2 kompleksi lõhustumist ja suurendab R- ja C-subühikute afiinsust ning moodustub PKA inaktiivne vorm.
Fosforüülitud ensüümid ja valgud fosfoproteiini fosfataas lähevad defosforüülitud vormi, muutub nende konformatsioon, aktiivsus ja protsesside kiirus, milles need ensüümid osalevad. Selle tulemusena naaseb süsteem algsesse olekusse ja on hormooni retseptoriga interaktsiooni korral uuesti aktiveerimiseks valmis. Seega on tagatud hormoonisisalduse vastavus veres ja sihtrakkude vastuse intensiivsus.
3. Adenülaattsüklaasi süsteemi osalemine geeniekspressiooni reguleerimises. Paljud valkhormoonid: glükagoon, vasopressiin, paratüreoidhormoon jne, mis edastavad oma signaali läbi adenülaattsüklaasi süsteemi, ei saa mitte ainult põhjustada reaktsioonide kiiruse muutust rakus juba olemasolevate ensüümide fosforüülimise teel, vaid ka suurendada või vähendada. nende arvu, reguleerides geeniekspressiooni (joonis 4.12). Aktiivne proteiinkinaas A võib siseneda tuuma ja fosforüülida transkriptsioonifaktorit (CREB). Fosfori liitumine
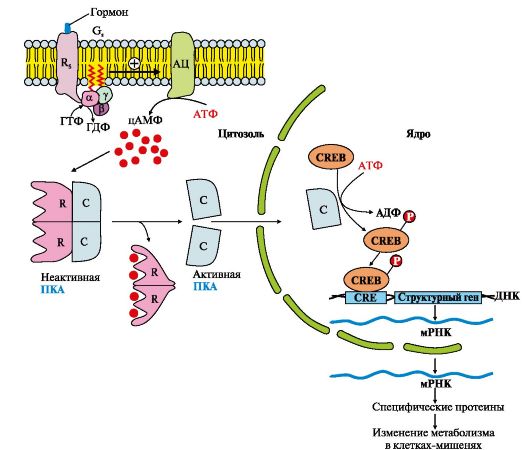 Riis. 4.12. Adenülaattsüklaasi rada, mis viib spetsiifiliste geenide ekspressioonini
Riis. 4.12. Adenülaattsüklaasi rada, mis viib spetsiifiliste geenide ekspressioonini
Jääk suurendab transkriptsioonifaktori (CREB-(P) afiinsust DNA-CRE regulatoorse tsooni spetsiifilise järjestuse suhtes (cAMP-response element) ja stimuleerib teatud valgugeenide ekspressiooni.
Sünteesitud valgud võivad olla ensüümid, mille koguse suurenemine suurendab ainevahetusprotsesside reaktsioonide kiirust, või membraanikandjad, mis tagavad teatud ioonide, vee või muude ainete rakku sisenemise või väljumise.

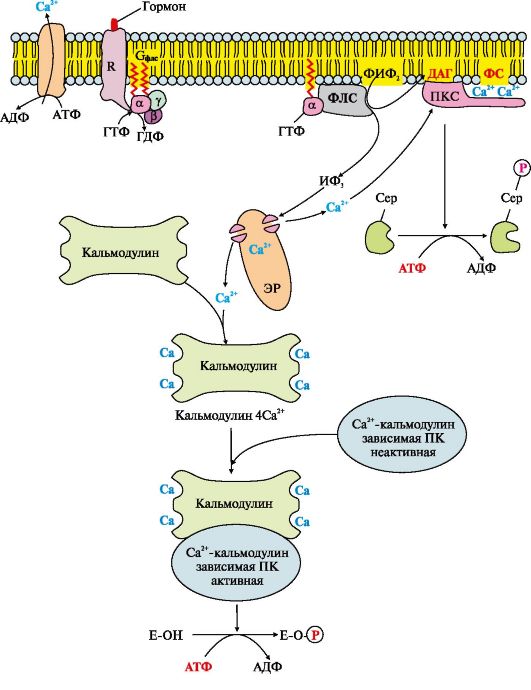 Riis. 4.13. Inositoolfosfaadi süsteem
Riis. 4.13. Inositoolfosfaadi süsteem
Süsteemi tööd tagavad valgud: kalmoduliin, ensüüm proteiinkinaas C, Ca 2 + -kalmoduliinist sõltuvad proteiinkinaasid, endoplasmaatilise retikulumi membraani reguleeritud Ca 2 + kanalid, raku ja mitokondri membraanide Ca 2 + -ATPaas .
Inositoolfosfaadi süsteemi primaarse messenger-signaali ülekande sündmuste jada
Inositoolfosfaatsüsteemi aktivaatori seondumine retseptoriga (R) viib selle konformatsiooni muutumiseni. Retseptori afiinsus Gf ls valgu suhtes suureneb. Primaarse messenger-retseptori kompleksi sidumine Gf ls-GDP-ga vähendab af ls-subühiku afiinsust SKT suhtes ja suurendab afiinsust GTP suhtes. Aktiivses kohas asendatakse SKT af ls allüksus GTP-ga. See põhjustab muutuse af ls subühiku konformatsioonis ja afiinsuse vähenemist βγ subühikute suhtes ning toimub Gf ls valgu dissotsiatsioon. Eraldunud subühik af ls-GTP liigub külgsuunas üle membraani ensüümi poole fosfolipaas C.
Ahls-GTP interaktsioon fosfolipaasi C sidumiskohaga muudab ensüümi konformatsiooni ja aktiivsust, suurendab rakumembraani fosfolipiidi, fosfatidüülinositool-4,5-bisfosfaadi (FIF 2) hüdrolüüsi kiirust (joonis 4.14).
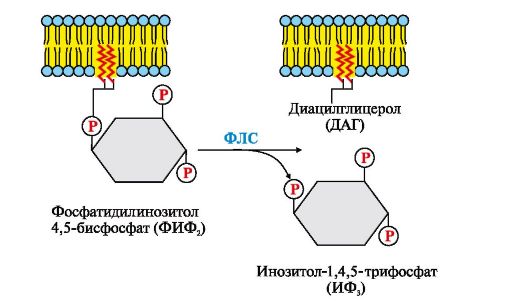 Riis. 4.14. Fosfatidüülinositool-4,5-bisfosfaadi (FIF 2) hüdrolüüs
Riis. 4.14. Fosfatidüülinositool-4,5-bisfosfaadi (FIF 2) hüdrolüüs
Reaktsiooni käigus moodustub kaks produkti - hormonaalse signaali sekundaarsed edastajad (sekundaarsed sõnumitoojad): diatsüülglütserool, mis jääb membraani ja osaleb proteiinkinaas C ensüümi aktiveerimises, ja inositool-1,4,5-trifosfaat. (IF 3), mis, olles hüdrofiilne ühend, läheb tsütosooli. Seega on rakuretseptori poolt vastuvõetud signaal kaheharuline. IP 3 seondub endoplasmaatilise retikulumi (E) membraani Ca 2+ kanali spetsiifiliste keskustega, mis viib valgu konformatsiooni muutumiseni ja Ca 2+ kanali avanemiseni. Kuna kaltsiumi kontsentratsioon ER-s on umbes 3-4 suurusjärku kõrgem kui tsütosoolis, siis pärast Ca 2+ kanali avanemist siseneb see tsütosooli mööda kontsentratsioonigradienti. IF 3 puudumisel tsütosoolis on kanal suletud.
Kõikide rakkude tsütosool sisaldab väikest valku nimega kalmoduliin, millel on neli Ca 2+ sidumissaiti. Suureneva kontsentratsiooniga
kaltsium, seondub see aktiivselt kalmoduliiniga, moodustades kompleksse 4Са 2+ -kalmoduliini. See kompleks interakteerub Ca 2+ -kalmoduliinist sõltuvate proteiinkinaaside ja teiste ensüümidega ning suurendab nende aktiivsust. Aktiveeritud Ca 2+-kalmoduliinist sõltuv proteiinkinaas fosforüleerib teatud valgud ja ensüümid, mille tulemusena muutub nende aktiivsus ja nende osalemise metaboolsete protsesside kiirus.
Ca 2+ kontsentratsiooni suurendamine raku tsütosoolis suurendab Ca 2+ interaktsiooni kiirust inaktiivse tsütosoolse ensüümiga proteiinkinaas C (PKC). PKC seondumine kaltsiumiioonidega stimuleerib valgu liikumist plasmamembraanile ja võimaldab ensüümil suhelda membraani fosfatidüülseriini (PS) molekulide negatiivselt laetud "peadega". Diatsüülglütserool, mis hõivab proteiinkinaas C spetsiifilisi kohti, suurendab veelgi selle afiinsust kaltsiumiioonide suhtes. Membraani siseküljel moodustub PKC aktiivne vorm (PKC? Ca2+? PS? DAG), mis fosforüleerib spetsiifilisi ensüüme.
IF-süsteemi aktiveerumine on lühiajaline ja pärast raku reageerimist stiimulile inaktiveeritakse fosfolipaas C, proteiinkinaas C ja Ca2+-kalmoduliinist sõltuvad ensüümid. af ls - GTP ja fosfolipaas C kompleksis olev allüksus omab ensümaatilise (GTP-fosfataasi) aktiivsust, hüdrolüüsib GTP-d. SKT-ga seotud af ls subühik kaotab oma afiinsuse fosfolipaasi C suhtes ja naaseb oma algsesse passiivsesse olekusse, s.t. sisaldub αβγ-SKT kompleksis Gf ls-valk).
Af ls-GDF eraldamine fosfolipaas C-st inaktiveerib ensüümi ja FIF 2 hüdrolüüs peatub. Ca 2+ kontsentratsiooni suurenemine tsütosoolis aktiveerib endoplasmaatilise retikulumi, tsütoplasmaatilise membraani, Ca 2+ -ATPaasi, mis "pumpab" raku tsütosoolist välja Ca 2 +. Selles protsessis osalevad ka Na+/Ca 2+- ja H+/Ca 2+-kandjad, mis toimivad aktiivse antiport põhimõttel. Ca 2+ kontsentratsiooni langus põhjustab Ca 2+ -kalmoduliinist sõltuvate ensüümide dissotsiatsiooni ja inaktiveerumist, samuti proteiinkinaas C afiinsuse kaotust membraani lipiidide suhtes ja selle aktiivsuse vähenemist.
Süsteemi aktiveerimise tulemusena moodustunud IP 3 ja DAG võivad taas üksteisega suhelda ja muutuda fosfatidüülinositool-4,5-bisfosfaadiks.
Fosforüülitud ensüümid ja valgud fosfoproteiinfosfataasi toimel muutuvad defosforüülitud vormiks, nende konformatsioon ja aktiivsus muutuvad.
5. Katalüütilised retseptorid. Katalüütilised retseptorid on ensüümid. Nende ensüümide aktivaatorid võivad olla hormoonid, kasvufaktorid, tsütokiinid. Aktiivsel kujul fosforüleerivad retseptor-ensüümid türosiini -OH rühmades spetsiifilisi valke, mistõttu neid nimetatakse türosiini proteiinkinaasideks (joonis 4.15). Spetsiaalsete mehhanismide kaudu saab katalüütilise retseptori vastuvõetud signaali edastada tuuma, kus see stimuleerib või pärsib teatud geenide ekspressiooni.
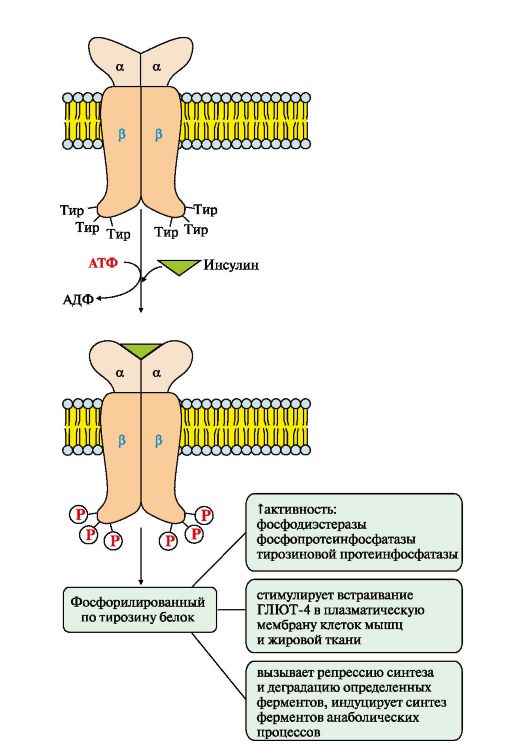 Riis. 4.15. Insuliini retseptori aktiveerimine.
Riis. 4.15. Insuliini retseptori aktiveerimine.
Fosfoproteiini fosfataas defosforüleerib spetsiifilisi fosfoproteiine.
Fosfodiesteraas muudab cAMP AMP-ks ja cGMP GMP-ks.
GLUT 4 – glükoosi transporterid insuliinist sõltuvates kudedes.
Türosiinvalgu fosfataas defosforüülib retseptori β-subühiku
insuliini
Katalüütilise retseptori näide on insuliini retseptor, mis koosneb kahest a- ja kahest β-subühikust. a-subühikud paiknevad rakumembraani välispinnal, β-subühikud tungivad läbi membraani kaksikkihi. Insuliini sidumissaidi moodustavad α-subühikute N-terminaalsed domeenid. Retseptori katalüütiline keskus asub β-subühikute rakusisestel domeenidel. Retseptori tsütosoolses osas on mitu türosiinijääki, mida saab fosforüülida ja defosforüülida.
Insuliini kinnitumine a-subühikute moodustatud sidumissaidile põhjustab retseptoris kooperatiivseid konformatsioonilisi muutusi. β-subühikutel on türosiinkinaasi aktiivsus ja need katalüüsivad transautofosforüülimist (esimene β-subühik fosforüülib teise β-subühiku ja vastupidi) mitme türosiini jäägi juures. Fosforüülimine põhjustab muutusi ensüümi (Tyr-PA) laengus, konformatsioonis ja substraadi spetsiifilisuses. Türosiin-PK fosforüülib teatud rakuvalke, mida nimetatakse insuliiniretseptori substraatideks. Need valgud on omakorda seotud fosforüülimisreaktsioonide kaskaadi aktiveerimisega:
• fosfoproteiini fosfataas(FPF), mis defosforüleerib spetsiifilisi fosfoproteiine;
• fosfodiesteraas, mis muudab cAMP AMP-ks ja cGMP GMP-ks;
• GLUT 4- glükoosikandjad insuliinsõltuvates kudedes, seetõttu suureneb glükoosi omastamine lihas- ja rasvkoe rakkudesse;
• türosiinvalgu fosfataas mis defosforüleerib insuliiniretseptori β-subühikut;
• tuuma reguleerivad valgud, transkriptsioonifaktorid, suurendada või vähendada teatud ensüümide geeniekspressiooni.
Efekti rakendamine kasvufaktorid saab läbi viia katalüütiliste retseptorite abil, mis koosnevad ühest polüpeptiidahelast, kuid moodustavad primaarse messengeri seondumisel dimeere. Kõikidel seda tüüpi retseptoritel on ekstratsellulaarne glükosüülitud domeen, transmembraanne (a-heeliks) ja tsütoplasmaatiline domeen, mis on võimeline aktiveerimisel avaldama proteiinkinaasi aktiivsust.
Dimerisatsioon aitab kaasa nende katalüütiliste intratsellulaarsete domeenide aktiveerimisele, mis teostavad seriini, treoniini või türosiini aminohappejääkide juures transautofosforüülimist. Fosforijääkide kinnitumine viib spetsiifiliste tsütosoolsete valkude seondumiskohtade moodustumiseni retseptoris ja proteiinkinaasi signaaliülekande kaskaadi aktiveerumiseni (joonis 4.16).
Primaarsete sõnumitoojate (kasvufaktorite) signaaliülekande sündmuste jada Ras- ja Raf-valkude osalusel.
Retseptori (R) seondumine kasvufaktoriga (GF) viib selle dimeriseerumiseni ja transautofosforüülimiseni. Fosforüülitud retseptor omandab afiinsuse Grb2 valgu suhtes. Moodustunud FR*R*Grb2 kompleks interakteerub tsütosoolse SOS-valguga. SOS konformatsiooni muutus
tagab selle interaktsiooni ankurdatud Ras-GDF membraanivalguga. FRaRgGrb2SOSRas-GDP kompleksi moodustumine vähendab Ras valgu afiinsust GDP suhtes ja suurendab afiinsust GTP suhtes.
GDP asendamine GTP-ga muudab Ras-valgu konformatsiooni, mis vabaneb kompleksist ja interakteerub membraanipiirkonnas Raf-valguga. Ras-GTP-Raf kompleksil on proteiinkinaasi aktiivsus ja see fosforüülib MEK kinaasi ensüümi. Aktiveeritud MEK kinaas omakorda fosforüülib MAP kinaasi treoniini ja türosiini juures.
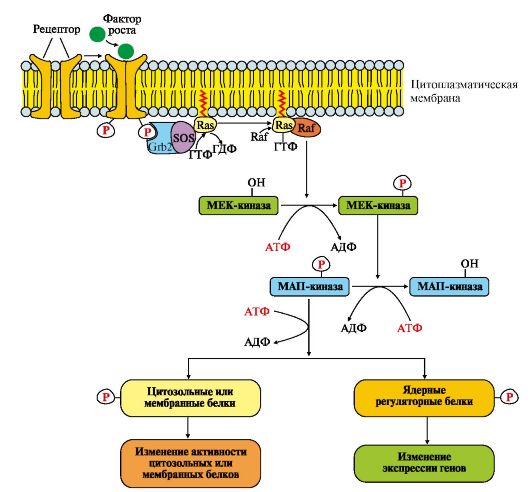 Joon.4.16. MAP kinaasi kaskaad.
Joon.4.16. MAP kinaasi kaskaad.
Seda tüüpi retseptoritel on epidermaalne kasvufaktor (EGF), närvikasvufaktor (NGF) ja muud kasvufaktorid.
Grb2 – valk, mis interakteerub kasvufaktori retseptoriga (kasvuretseptorit siduv valk); SOS (GEF) - GDP-GTP vahetusfaktor (guaniini nukleotiidi vahetusfaktor); Ras - G-valk (guanidiintrifosfataas); Raf-kinaas - aktiivsel kujul - fosforüüliv MEK-kinaas; MEK kinaas - MAP kinaasi kinaas; MAP kinaas – mitogeeniga aktiveeritud proteiinkinaas (mitogeeniga aktiveeritud proteiinkinaas)
-PO 3 2 - rühma kinnitumine MAP kinaasi aminohapete radikaalidele muudab selle laengut, konformatsiooni ja aktiivsust. Ensüüm fosforüülib membraanide, tsütosooli ja tuuma spetsiifilisi valke seriiniks ja treoniiniks.
Muutused nende valkude aktiivsuses mõjutavad ainevahetusprotsesside kiirust, membraani translokaaside toimimist ja sihtrakkude mitootilist aktiivsust.
Retseptorid koos guanülaattsüklaasi aktiivsus Neid nimetatakse ka katalüütilisteks retseptoriteks. Guanülaadi tsüklaas katalüüsib cGMP moodustumist GTP-st, mis on rakusisese signaaliülekande üks olulisi sõnumitoojaid (vahendajaid) (joonis 4.17).
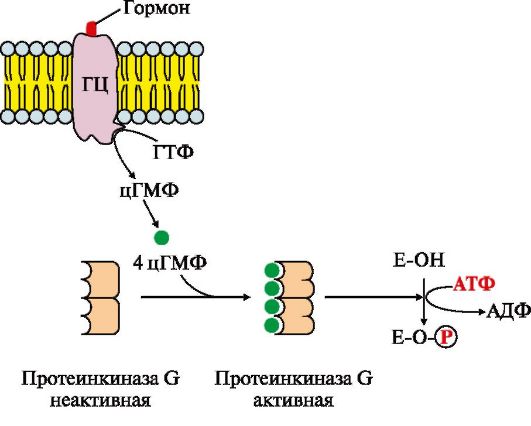 Riis. 4.17. Membraani guanülaattsüklaasi aktiivsuse reguleerimine.
Riis. 4.17. Membraani guanülaattsüklaasi aktiivsuse reguleerimine.
Membraaniga seotud guanülaattsüklaas (GC) on transmembraanne glükoproteiin. Signaalmolekuli sidumiskeskus asub ekstratsellulaarsel domeenil, guanülaattsüklaasi intratsellulaarne domeen avaldab aktiveerimise tulemusena katalüütilist aktiivsust
Primaarse messengeri kinnitumine retseptorile aktiveerib guanülaattsüklaasi, mis katalüüsib GTP konversiooni tsükliliseks guanosiin-3,5'-monofosfaadiks (cGMP), teiseks sõnumitoojaks. CGMP kontsentratsioon rakus suureneb. cGMP molekulid võivad pöörduvalt kinnituda proteiinkinaas G (PKG5) regulatoorsete keskustega, mis koosneb kahest subühikust. Neli cGMP molekuli muudavad ensüümi konformatsiooni ja aktiivsust. Aktiivne proteiinkinaas G katalüüsib teatud valkude ja ensüümide fosforüülimist raku tsütosoolis. Üks proteiinkinaas G peamisi sõnumitoojaid on kodade natriureetiline faktor (ANF), mis reguleerib vedeliku homöostaasi kehas.
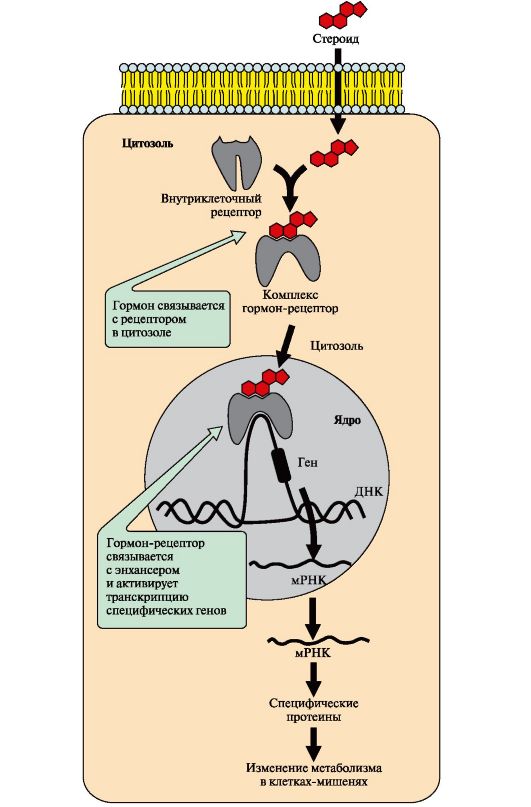
6. Signaali edastamine intratsellulaarsete retseptorite abil. Keemiliselt hüdrofoobsed hormoonid (steroidhormoonid ja türoksiin) võivad difundeeruda läbi membraanide, mistõttu nende retseptorid paiknevad tsütosoolis või raku tuumas.
Tsütosoolsed retseptorid on seotud chaperone valguga, mis takistab retseptori enneaegset aktivatsiooni. Steroid- ja kilpnäärmehormoonide tuuma- ja tsütosoolretseptorid sisaldavad DNA-d siduvat domeeni, mis tagab hormoon-retseptori kompleksi koostoime tuumas oleva DNA regulatoorsete piirkondadega ja transkriptsioonikiiruse muutused.
Sündmuste jada, mis põhjustab transkriptsioonikiiruse muutumise
Hormoon läbib rakumembraani lipiidide kaksikkihti. Tsütosoolis või tuumas interakteerub hormoon retseptoriga. Hormoon-retseptori kompleks siseneb tuuma ja kinnitub DNA regulatoorse nukleotiidjärjestuse külge - võimendaja(joon. 4.18) või summuti. RNA polümeraasi promootori kättesaadavus suureneb interaktsioonil võimendajaga või väheneb interaktsioonil summutiga. Vastavalt sellele suureneb või väheneb teatud struktuursete geenide transkriptsiooni kiirus. Küpsed mRNA-d vabanevad tuumast. Teatud valkude translatsiooni kiirus suureneb või väheneb. Muutub raku ainevahetust ja funktsionaalset seisundit mõjutavate valkude hulk.
Igas rakus on erinevatesse signaalimuundurisüsteemidesse kaasatud retseptorid, mis muudavad kõik välised signaalid rakusiseseks. Konkreetse esmase sõnumitooja retseptorite arv võib varieeruda 500-st üle 100 000-ni raku kohta. Need asuvad membraanil üksteisest eemal või on koondunud selle teatud piirkondadesse.
 Riis. 4.18. Signaali ülekanne rakusisestele retseptoritele
Riis. 4.18. Signaali ülekanne rakusisestele retseptoritele
b) valige tabelist lipiidid, mis on seotud:
1. Proteiinkinaas C aktiveerimine
2. DAG moodustumise reaktsioonid fosfolipaasi C toimel
3. Närvikiudude müeliinkestade moodustumine
c) kirjutage lõikes 2 valitud lipiidi hüdrolüüsireaktsioon;
d) näidata, milline hüdrolüüsiproduktidest osaleb endoplasmaatilise retikulumi Ca 2 + kanali reguleerimises.
2. Vali õiged vastused.
Kandjavalkude konformatsioonilist labiilsust võivad mõjutada:
B. Elektrilise potentsiaali muutus läbi membraani
B. Spetsiifiliste molekulide kinnitumine D. Kahekihiliste lipiidide rasvhapete koostis E. Transporditava aine kogus
3. Määra sobivus:
A. ER kaltsiumikanal B. Ca 2 +-ATPaas
D. Ka +-sõltuv kandja Ca 2 + D. N +, K + -ATPaas
1. Kannab Na+ piki kontsentratsioonigradienti
2. Toimib hõlbustatud difusiooni mehhanismi järgi
3. Kannab Na+ vastu kontsentratsioonigradienti
4. Viige laud üle. 4.2. märkmik ja täitke see.
Tabel 4.2. Adenülaattsüklaasi ja inositoolfosfaadi süsteemid
Struktuur ja tööetapid | Adenülaattsüklaasi süsteem | Inositoolfosfaadi süsteem |
Süsteemi esmase sõnumitooja näide | ||
Integreeritud rakumembraani valk, mis interakteerub komplementaarselt primaarse sõnumitoojaga | ||
Signaalensüümi aktiveeriv valk | ||
Ensüümsüsteem, mis moodustab sekundaarse(e) sõnumikandja(d) | ||
Süsteemi sekundaarsed sõnumitoojad | ||
Süsteemi tsütosoolne (e) ensüüm(id), mis interakteeruvad (e) teise sõnumitoojaga | ||
Metaboolsete radade ensüümide aktiivsuse reguleerimise mehhanism (selles süsteemis). | ||
Mehhanismid sekundaarsete sõnumitoojate kontsentratsiooni vähendamiseks sihtrakus | ||
Signaalsüsteemi membraani ensüümi aktiivsuse vähenemise põhjus |
ÜLESANDED ENESEKOHTAMISEKS
1. Määra sobivus:
A. Passiivne sümptom B. Passiivne antiport
B. Endotsütoos D. Eksotsütoos
D. Esmane aktiivne transport
1. Aine transport rakku toimub koos plasmamembraani osaga
2. Kontsentratsioonigradienti mööda läheb rakku samaaegselt kaks erinevat ainet
3. Ainete transport läheb vastuollu kontsentratsioonigradiendiga
2. Vali õige vastus.
ag-GTP-ga seotud G-valgu subühik aktiveerib:
A. Retseptor
B. Proteiini kinaas A
B. Fosfodiesteraas D. Adenülaattsüklaas E. Proteiini kinaas C
3. Määra vaste.
Funktsioon:
A. Reguleerib katalüütilise retseptori aktiivsust B. Aktiveerib fosfolipaasi C
B. Muudab proteiinkinaasi A aktiivseks vormiks
D. Suurendab Ca 2+ kontsentratsiooni raku tsütosoolis E. Aktiveerib proteiinkinaasi C
Teine sõnumitooja:
4. Määra vaste.
Toimimine:
A. Võimalik külgdifusiooniks membraani kaksikkihis
B. Koos esmase sõnumitoojaga liitub see võimendajaga
B. Näitab ensümaatilist aktiivsust esmase sõnumitoojaga suheldes
G. Võib interakteeruda G-valguga
D. Suhtleb signaali edastamise ajal fosfolipaas C-ga Retseptor:
1. Insuliin
2. Adrenaliin
3. Steroidhormoon
5. Täitke "ahela" ülesanne:
a) peptiidhormoonid interakteeruvad retseptoritega:
A. Raku tsütosoolis
B. Sihtrakumembraanide integraalsed valgud
B. Raku tuumas
G. Kovalentselt seotud FIF 2-ga
b) sellise retseptori koostoime hormooniga põhjustab kontsentratsiooni suurenemist rakus:
A. Hormoon
B. Vahemetaboliidid
B. Teised sõnumitoojad D. Tuumavalgud
sisse) Need molekulid võivad olla:
A. MÄRGIST B. GTP
B. FIF 2 D. cAMP
G) nad aktiveerivad:
A. Adenülaattsüklaas
B. Ca 2+ -sõltuv kalmoduliin
B. Proteiini kinaas A D. Fosfolipaas C
e) see ensüüm muudab metaboolsete protsesside kiirust rakus:
A. Ca 2 + kontsentratsiooni suurendamine tsütosoolis B. Reguleerivate ensüümide fosforüülimine
B. Protenfosfataasi aktiveerimine
D. Muutused regulatoorsete valkude geenide ekspressioonis
6. Täitke "ahela" ülesanne:
a) kasvufaktori (GF) kinnitumine retseptorile (R) põhjustab:
A. Muutused FR-R kompleksi lokaliseerimises
B. Retseptori dimerisatsioon ja transautofosforüülimine
B. Muutus retseptori konformatsioonis ja kinnitumine Gs valguga D. FR-R kompleksi liikumine
b) sellised muutused retseptori struktuuris suurendavad selle afiinsust membraani pinnavalgu suhtes:
B. Raf G. Grb2
sisse) see interaktsioon suurendab tsütosoolse valgukompleksi külge kinnitumise tõenäosust:
A. Kalmodulina B. Ras
B. PCS D. SOS
G) mis suurendab kompleksi komplementaarsust "ankurdatud" valguga:
e) "ankurdatud" valgu konformatsiooni muutus vähendab selle afiinsust:
A. CAMP B. GTP
B. GDF G. ATP
e) see aine asendatakse järgmisega:
A. GDF B. AMP
B. cGMP D. GTP
ja) nukleotiidi lisamine soodustab "ankurdatud" valgu koostoimet:
A. PKA B. Kalmoduliin
h) See valk on osa kompleksist, mis fosforüülib:
A. MEK kinaas B. Proteiini kinaas C
B. Proteiini kinaas A D. MAP kinaas
ja) See ensüüm omakorda aktiveerib:
A. MEK kinaas B. Proteiini kinaas G
B. Raf proteiin D. MAP kinaas
j) valgu fosforüülimine suurendab selle afiinsust:
A. SOS- ja Raf-valgud B. Tuuma reguleerivad valgud B. Kalmoduliin D. Tuumaretseptorid
k) nende valkude aktiveerimine põhjustab:
A. GTP defosforüülimine Ras-valgu aktiivses keskuses B. Retseptori vähenenud afiinsus kasvufaktori suhtes
B. Maatriksi biosünteesi kiiruse suurenemine D. SOS-Grb2 kompleksi dissotsiatsioon
m) selle tulemusena:
A. SOS valk vabaneb retseptorist
B. Toimub retseptori protomeeride (R) dissotsiatsioon
B. Ras-valk eraldub Raf-valgust
D. Sihtraku proliferatiivne aktiivsus suureneb.
VASTUSTE STANDARDID "ENESEKOHTAMISEKS"
1. 1-B, 2-A, 3-D
3. 1-B, 2-D, 3-G
4. 1-C, 2-D, 3-B
5. a) B, b) C, c) D, d) C, e) B
6. a) B, b) D, c) D, d) A, e) B, f) D, g) D, h) A, i) D, j) C, l) C, m) D
PÕHIMÕISTED JA MÕISTED
1. Membraanide ehitus ja funktsioonid
2. Ainete transport läbi membraanide
3. Membraanivalkude ehituslikud iseärasused
4. Transmembraansed signaalisüsteemid (adenülaattsüklaas, inositoolfosfaat, guanülaattsüklaas, katalüütilised ja intratsellulaarsed retseptorid)
5. Esmased sõnumitoojad
6. Teisesed sõnumitoojad (vahendajad)
AUDIITSIOONITÖÖD ÜLESANDED
1. Vaata joon. 4.19 ja täitke järgmised ülesanded:
a) nimetada transpordiliik;
b) määrake sündmuste järjekord:
A. Cl - lahkub rakust piki kontsentratsioonigradienti
B. Proteiini kinaas A fosforüülib kanali R-subühiku
B. R-subühiku konformatsiooni muutused
D. Membraanivalgus esinevad kooperatiivsed konformatsioonilised muutused
D. Adenülaattsüklaasi süsteem on aktiveeritud
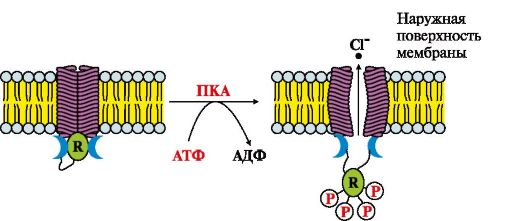 Riis. 4.19. Soole endoteeli C1-kanali toimimine.
Riis. 4.19. Soole endoteeli C1-kanali toimimine.
R on reguleeriv valk, mis muundatakse proteiinkinaas A (PKA) toimel fosforüülitud vormiks.
c) võrdleb endoplasmaatilise retikulummembraani Ca 2+ kanali ja soole endoteeliraku Cl - kanali talitlust, täites tabeli. 4.3.
Tabel 4.3. Kanalite toimimise reguleerimise viisid
Probleeme lahendama
1. Südamelihase kontraktsioon aktiveerib Ca 2 +, mille sisaldus raku tsütosoolis suureneb tsütoplasmaatilise membraani cAMP-sõltuvate kandjate toimimise tõttu. Omakorda reguleerivad cAMP kontsentratsiooni rakkudes kaks signaalmolekuli - adrenaliin ja atsetüülkoliin. Lisaks on teada, et β2-adrenergiliste retseptoritega interakteeruv adrenaliin suurendab cAMP kontsentratsiooni müokardi rakkudes ja stimuleerib südame väljundit ning atsetüülkoliin, interakteerudes M 2 -kolinergiliste retseptoritega, vähendab cAMP taset ja müokardi kontraktiilsust. Selgitage, miks kaks peamist sõnumitoojat, kasutades sama signaaliülekandesüsteemi, kutsuvad esile erineva rakulise vastuse. Selle jaoks:
a) esitab adrenaliini ja atsetüülkoliini signaaliülekande skeemi;
b) näitavad nende sõnumitoojate signaalikaskaadide erinevust.
2. Atsetüülkoliin, interakteerudes süljenäärmete M 3 -kolinergiliste retseptoritega, stimuleerib Ca 2+ vabanemist ER-st. Ca 2+ kontsentratsiooni tõus tsütosoolis tagab sekretoorsete graanulite eksotsütoosi ning elektrolüütide ja väikese koguse valkude vabanemise süljejuhasse. Selgitage, kuidas reguleeritakse ER-i Ca 2+ kanaleid. Selle jaoks:
a) nimetage ER Ca 2+ kanalite avamist võimaldav teine sõnumitooja;
b) kirjutada reaktsioon teise sõnumitooja tekkeks;
c) esitab atsetüülkoliini transmembraanse signaaliülekande skeemi, mille aktiveerimise käigus saab regulaatorligand Ca 2+ -
3. Insuliiniretseptorite uurijad on tuvastanud olulise muutuse valgu, ühe insuliiniretseptori substraadi geenis. Kuidas mõjutab häire selle valgu struktuuris insuliini signaalisüsteemi toimimist? Küsimusele vastamiseks:
a) esitage insuliini transmembraanse signaalimise diagramm;
b) nimetada sihtrakkudes insuliini aktiveerivad valgud ja ensüümid, näidata nende funktsioon.
4. Ras-valk on tsütoplasmaatilises membraanis "ankurdatud" valk. "Ankru" funktsiooni täidab farnesüüli H 3 C-(CH 3) C \u003d CH-CH 2 - [CH 2 - (CH 3) C \u003d CH-CH 2] 2 15-süsiniku jääk -, mis kinnitub valgu külge ensüümi farnesüültransferaas kaudu translatsioonijärgse modifikatsiooni käigus. Praegu läbivad selle ensüümi inhibiitorid kliinilised uuringud.
Miks nende ravimite kasutamine kahjustab kasvufaktori signaaliülekannet? Vastuseks:
a) esitab Ras valke hõlmava signaaliülekande skeemi;
b) selgitab Ras-valkude funktsiooni ja nende atsüülimise ebaõnnestumise tagajärgi;
Loodus on loonud palju organisme ja rakke, kuid vaatamata sellele on bioloogiliste membraanide struktuur ja enamik funktsioone samad, mis võimaldab kaaluda nende struktuuri ja uurida nende võtmeomadusi, olemata seotud teatud tüüpi rakuga.
Mis on membraan?
Membraanid on kaitseelement, mis on iga elusorganismi raku lahutamatu osa.
Kõigi planeedi elusorganismide struktuurne ja funktsionaalne üksus on rakk. Tema elutähtis tegevus on lahutamatult seotud keskkonnaga, millega ta vahetab energiat, teavet, ainet. Seega tuleb raku toimimiseks vajalik toitumisenergia väljastpoolt ja kulub selle erinevate funktsioonide elluviimiseks.
Elusorganismi kõige lihtsama struktuuriüksuse ehitus: organellmembraan, mitmesugused kandmised. Seda ümbritseb membraan, mille sees asuvad tuum ja kõik organellid. Need on mitokondrid, lüsosoomid, ribosoomid, endoplasmaatiline retikulum. Igal konstruktsioonielemendil on oma membraan.
Roll raku elus
Bioloogiline membraan mängib elementaarse elusüsteemi ülesehituses ja toimimises kulmineerivat rolli. Organismiks võib õigusega nimetada vaid rakku, mida ümbritseb kaitsekesta. Membraani olemasolu tõttu toimub ka selline protsess nagu ainevahetus. Kui selle struktuurset terviklikkust rikutakse, muutub see organismi kui terviku funktsionaalses seisundis.
Rakumembraan ja selle funktsioonid
See eraldab raku tsütoplasma väliskeskkonnast või membraanist. Rakumembraan tagab spetsiifiliste funktsioonide korrektse täitmise, rakkudevaheliste kontaktide ja immuunilmingute spetsiifika ning toetab transmembraanset elektripotentsiaali erinevust. See sisaldab retseptoreid, mis suudavad tajuda keemilisi signaale – hormoone, vahendajaid ja muid bioloogiliselt aktiivseid komponente. Need retseptorid annavad sellele veel ühe võime – muuta raku metaboolset aktiivsust.
Membraani funktsioonid:
1. Ainete aktiivne ülekanne.
2. Ainete passiivne ülekanne:
2.1. Difusioon on lihtne.
2.2. transportida läbi pooride.
2.3. Transport toimub kandja difusiooni teel koos membraanainega või aine edasikandmisega mööda kandja molekulaarahelat.
3. Mitteelektrolüütide ülekanne lihtsa ja hõlbustatud difusiooni tõttu.
4. Ioonide aktiivne transport.
Rakumembraani struktuur
Rakumembraani komponendid on lipiidid ja valgud.
Lipiidid: fosfolipiidid, fosfatidüületanoolamiin, sfingomüeliin, fosfatidüülinositool ja fosfatidüülseriin, glükolipiidid. Lipiidide osakaal on 40-90%.
Valgud: perifeersed, integraalsed (glükoproteiinid), spektriin, aktiin, tsütoskelett.
Peamine struktuurielement on fosfolipiidimolekulide topeltkiht.
Katusemembraan: määratlus ja tüpoloogia
Natuke statistikat. Vene Föderatsiooni territooriumil on membraani kasutatud katusekattematerjalina mitte nii kaua aega tagasi. Membraankatuste osakaal pehmete katuseplaatide koguarvust on vaid 1,5%. Bituumen- ja mastikskatused on Venemaal laiemalt levinud. Kuid Lääne-Euroopas moodustavad membraankatused 87%. Erinevus on tuntav.
Reeglina sobib membraan katusekatte põhimaterjalina ideaalselt lamekatuste jaoks. Neile, kellel on suur eelarvamus, sobib see vähem.
Siseturu membraankatuste tootmis- ja müügimahud on positiivse kasvutrendiga. Miks? Põhjused on enam kui selged:
- Kasutusiga on umbes 60 aastat. Kujutage ette, ainult tootja poolt määratud kasutusgarantii ulatub 20 aastani.
- Paigaldamise lihtsus. Võrdluseks: bituumenkatuse paigaldamine võtab 1,5 korda rohkem aega kui membraanpõranda paigaldamine.
- Hooldus- ja remonditööde lihtsus.
Katusemembraanide paksus võib olla 0,8-2 mm ja ühe ruutmeetri keskmine kaal on 1,3 kg.
Katusemembraanide omadused:
- elastsus;
- tugevus;
- vastupidavus ultraviolettkiirtele ja muudele agressoritele;
- külmakindlus;
- tulekindlus.
Katusemembraane on kolme tüüpi. Peamine klassifitseerimistunnus on polümeermaterjali tüüp, mis moodustab lõuendi aluse. Niisiis on katusemembraanid:
- EPDM rühma kuuluvad, on valmistatud polümeriseeritud etüleen-propüleen-dieeni monomeeri baasil, teisisõnu Eelised: kõrge tugevus, elastsus, veekindlus, keskkonnasõbralikkus, madal hind. Puudused: liimitehnoloogia lõuendite ühendamiseks spetsiaalse teibi abil, madala tugevusega liitekohad. Kasutusala: kasutatakse hüdroisolatsioonimaterjalina tunneli lagede, veeallikate, jäätmehoidlate, tehis- ja looduslike veehoidlate jms jaoks.
- PVC membraanid. Need on kestad, mille valmistamisel kasutatakse põhimaterjalina polüvinüülkloriidi. Eelised: vastupidavus ultraviolettkiirgusele, tulekindlus, membraanilehtede lai värvivalik. Puudused: madal vastupidavus bituumenmaterjalidele, õlidele, lahustitele; eraldab atmosfääri kahjulikke aineid; lõuendi värv tuhmub aja jooksul.
- TPO. Valmistatud termoplastilistest olefiinidest. Neid saab tugevdada ja tugevdamata. Esimesed on varustatud polüestervõrgu või klaaskiudlapiga. Eelised: keskkonnasõbralikkus, vastupidavus, kõrge elastsus, temperatuuritaluvus (nii kõrgel kui ka madalal temperatuuril), lõuendi õmbluste keevisliited. Puudused: kõrge hinnakategooria, tootjate puudumine siseturul.
Profileeritud membraan: omadused, funktsioonid ja eelised
Profileeritud membraanid on ehitusturu uuendus. Sellist membraani kasutatakse hüdroisolatsioonimaterjalina.

Valmistamisel kasutatud materjal on polüetüleen. Viimast on kahte tüüpi: kõrgsurvepolüetüleen (LDPE) ja madalrõhupolüetüleen (HDPE).
Indeks | ||
Tõmbetugevus (MPa) | ||
Tõmbe pikenemine (%) | ||
Tihedus (kg / m3) | ||
Survetugevus (MPa) | ||
Löögitugevus (sälguga) (KJ/ruutmeetri kohta) | ||
Paindemoodul (MPa) | ||
Kõvadus (MPa) | ||
Töötemperatuur (˚С) | -60 kuni +80 | -60 kuni +80 |
Päevane veeimavus (%) |
Kõrgsurve polüetüleenist valmistatud profiilmembraanil on spetsiaalne pind - õõnsad vistrikud. Nende koosseisude kõrgus võib varieeruda 7 kuni 20 mm. Membraani sisepind on sile. See võimaldab ehitusmaterjalide tõrgeteta painutamist.
Membraani üksikute sektsioonide kuju muutumine on välistatud, kuna rõhk jaotub kõigi samade eendite olemasolu tõttu ühtlaselt kogu selle alale. Geomembraani saab kasutada ventilatsiooni isolatsioonina. Sel juhul on tagatud hoonesisene vaba soojusvahetus.
Profileeritud membraanide eelised:
- suurenenud tugevus;
- kuumakindlus;
- keemilise ja bioloogilise mõju stabiilsus;
- pikk kasutusiga (üle 50 aasta);
- paigaldamise ja hooldamise lihtsus;
- taskukohane hind.
Profileeritud membraane on kolme tüüpi:
- ühe kihiga;
- kahekihilise lõuendiga = geotekstiil + drenaažimembraan;
- kolmekihilise lõuendiga = libe pind + geotekstiil + drenaažimembraan.
Ühekihilist profiilmembraani kasutatakse kõrge õhuniiskusega seinte peamise hüdroisolatsiooni, betooni ettevalmistamise, paigaldamise ja demonteerimise kaitsmiseks. Seina äravoolu varustamisel kasutatakse kahekihilist kaitset. Kolmest kihist koosnevat seda kasutatakse külmakindlal pinnasel ja sügaval pinnasel.
Drenaažimembraanide kasutusvaldkonnad
Profileeritud membraani kasutatakse järgmistes valdkondades:
- Põhiline vundamendi hüdroisolatsioon. Pakub usaldusväärset kaitset põhjavee, taimede juurestiku, pinnase vajumise ja mehaaniliste kahjustuste hävitava mõju eest.
- Vundamendi seina drenaaž. Neutraliseerib põhjavee, sademete mõju, viies need ära kuivendussüsteemidesse.
- Horisontaalne reservuaari tüüpi drenaaž - kaitse konstruktsioonilistest iseärasustest tingitud deformatsiooni eest.
- Betooni valmistamise analoog. Seda kasutatakse madala põhjavee tsoonis asuvate hoonete ehitustöödel juhtudel, kui kapillaarniiskuse eest kaitsmiseks kasutatakse horisontaalset hüdroisolatsiooni. Samuti kuuluvad profiilmembraani funktsioonide hulka tsemendipiima pinnasesse mitteläbilaskvus.
- Kõrge niiskustasemega seinapindade ventilatsioon. Seda saab paigaldada nii ruumi sise- kui ka välisküljele. Esimesel juhul aktiveeritakse õhuringlus ja teisel on tagatud optimaalne niiskus ja temperatuur.
- Kasutatud tagurpidi katus.
Super difusioonmembraan
Tegemist on uue põlvkonna materjaliga, mille põhieesmärk on kaitsta katusekonstruktsiooni elemente tuulenähtuste, sademete ja auru eest.

Kaitsematerjali tootmine põhineb lausriide, kvaliteetsete tihedate kiudude kasutamisel. Siseturul on populaarne kolmekihiline ja neljakihiline membraan. Ekspertide ja tarbijate ülevaated kinnitavad, et mida rohkem kihte on disainil, seda tugevamad on selle kaitsefunktsioonid ja seega ka ruumi kui terviku energiatõhusus.
Sõltuvalt katuse tüübist, selle konstruktsiooniomadustest, kliimatingimustest soovitavad tootjad eelistada üht või teist tüüpi difusioonmembraane. Niisiis, need on olemas keerukate ja lihtsate konstruktsioonidega viilkatuste jaoks, minimaalse kaldega viilkatuste jaoks, voldikkatuste jaoks jne.
Superdifusioonmembraan asetatakse otse soojusisolatsioonikihile, põrandakate laudadest. Tuulutuspilu pole vaja. Materjal kinnitatakse spetsiaalsete sulgude või terasnaeltega. Difusiooniplekkide servad on ühendatud.Tööd saab teha ka ekstreemsetes tingimustes: tugevate tuuleiilide korral jne.

Lisaks saab kõnealust katet kasutada ajutise katusekattena.
PVC membraanid: olemus ja eesmärk
PVC membraanid on polüvinüülkloriidist valmistatud katusematerjal, millel on elastsed omadused. Selline kaasaegne katusematerjal asendas täielikult bituumenrulli analoogid, millel on märkimisväärne puudus - vajadus süstemaatilise hoolduse ja remondi järele. Tänapäeval võimaldavad PVC-membraanide iseloomulikud omadused neid kasutada vanade lamekatuste remonditöödel. Neid kasutatakse ka uute katuste paigaldamisel.

Sellisest materjalist katust on lihtne kasutada ning selle paigaldamine on võimalik igat tüüpi pinnale, igal aastaajal ja igasuguste ilmastikutingimuste korral. PVC membraanil on järgmised omadused:
- tugevus;
- stabiilsus UV-kiirte, erinevat tüüpi sademete, punkt- ja pinnakoormusega kokkupuutel.
Tänu oma ainulaadsetele omadustele teenivad PVC membraanid teid ustavalt palju aastaid. Sellise katuse kasutusaeg on võrdne hoone enda kasutusajaga, samas kui valtskattematerjalid vajavad regulaarset remonti ja mõnel juhul isegi demonteerimist ja uue põranda paigaldamist.
PVC membraanlehed ühendatakse omavahel kuumhingamiskeevitusega, mille temperatuur jääb vahemikku 400-600 kraadi Celsiuse järgi. See ühendus on täielikult suletud.
PVC membraanide eelised
Nende eelised on ilmsed:
- katusesüsteemi paindlikkus, mis on kõige paremini kooskõlas ehitusprojektiga;
- vastupidav, õhutihe ühendusõmblus membraanilehtede vahel;
- ideaalne taluvus kliimamuutuste, ilmastikutingimuste, temperatuuri, niiskuse suhtes;
- suurenenud auru läbilaskvus, mis aitab kaasa katusealusesse ruumi kogunenud niiskuse aurustumisele;
- palju värvivalikuid;
- tulekustutusomadused;
- võime säilitada originaalseid omadusi ja välimust pikka aega;
- PVC membraan on absoluutselt keskkonnasõbralik materjal, mida kinnitavad vastavad sertifikaadid;
- paigaldusprotsess on mehhaniseeritud, nii et see ei võta palju aega;
- kasutusreeglid võimaldavad paigaldada erinevaid arhitektuurseid täiendusi otse PVC membraankatuse enda peale;
- ühekihiline stiil säästab teie raha;
- hoolduse ja remondi lihtsus.

Membraankangas
Membraankangas on tekstiilitööstusele tuntud juba pikka aega. Jalatsid ja riided on valmistatud sellisest materjalist: täiskasvanutele ja lastele. Membraan - membraankanga alus, mis on esitatud õhukese polümeerkile kujul ja millel on sellised omadused nagu veekindlus ja auru läbilaskvus. Selle materjali tootmiseks kaetakse see kile välimise ja sisemise kaitsekihiga. Nende struktuuri määrab membraan ise. Seda tehakse selleks, et säilitada kõik kasulikud omadused ka kahjustuste korral. Teisisõnu, membraanirõivad ei saa märjaks, kui nad puutuvad kokku lume või vihma näol sademetega, kuid juhivad samal ajal suurepäraselt auru kehast väliskeskkonda. See läbilaskvus võimaldab nahal hingata.
Kõike eelnevat arvestades võime järeldada, et just sellisest kangast valmivad ideaalsed talveriided. Membraan, mis on kanga põhjas, võib olla:
- pooridega;
- ilma poorideta;
- kombineeritud.

Teflon sisaldub paljude mikropooridega membraanide koostises. Selliste pooride mõõtmed ei küüni isegi veetilga mõõtmeteni, vaid on suuremad kui veemolekul, mis viitab veekindlusele ja võimele higi eemaldada.
Membraanid, millel pole poore, on tavaliselt valmistatud polüuretaanist. Nende sisemine kiht koondab kõik inimkeha higi-rasvaeritused ja surub need välja.
Kombineeritud membraani struktuur eeldab kahe kihi olemasolu: poorne ja sile. Sellel kangal on kõrged kvaliteediomadused ja see kestab palju aastaid.
Tänu neile eelistele on talvehooajal kandmiseks mõeldud membraankangast riided ja jalanõud vastupidavad, kuid kerged ning kaitsevad suurepäraselt külma, niiskuse ja tolmu eest. Need on lihtsalt asendamatud paljude aktiivsete talvise puhkuse, alpinismi jaoks.
bioloogilised membraanid.
Mõistet "membraan" (ladina keeles membrana – nahk, kile) hakati kasutama rohkem kui 100 aastat tagasi, tähistamaks rakupiiri, toimides ühelt poolt barjäärina raku sisu ja väliskeskkonna vahel, ja teiselt poolt poolläbilaskva vaheseinana, millest pääseb läbi vesi ja mõned ained. Kuid membraani funktsioonid ei ole ammendatud, kuna bioloogilised membraanid moodustavad raku struktuurilise korralduse aluse.
Membraani struktuur. Selle mudeli järgi on põhimembraaniks lipiidide kaksikkiht, milles molekulide hüdrofoobsed sabad on pööratud sissepoole ja hüdrofiilsed pead väljapoole. Lipiide esindavad fosfolipiidid - glütserooli või sfingosiini derivaadid. Valgud kinnituvad lipiidikihile. Integraalsed (transmembraansed) valgud tungivad läbi membraani ja on sellega kindlalt seotud; perifeersed ei tungi ja on membraaniga vähem kindlalt seotud. Membraanivalkude funktsioonid: membraanide struktuuri säilitamine, keskkonna signaalide vastuvõtmine ja muundamine. keskkond, teatud ainete transport, membraanidel toimuvate reaktsioonide katalüüs. membraani paksus on 6 kuni 10 nm.
Membraani omadused:
1. Sujuvus. Membraan ei ole jäik struktuur, suurem osa selle valkudest ja lipiididest võivad liikuda membraanide tasapinnal.
2. Asümmeetria. Nii valkude kui ka lipiidide välise ja sisemise kihi koostis on erinev. Lisaks on loomarakkude plasmamembraanidel väljastpoolt glükoproteiinide kiht (glükokalüks, mis täidab signaali- ja retseptori funktsioone ning on oluline ka rakkude ühendamisel kudedeks)
3. Polaarsus. Membraani väliskülg kannab positiivset laengut, sisemine aga negatiivset laengut.
4. Valikuline läbilaskvus. Elusrakkude membraanid läbivad lisaks veele ainult teatud molekule ja lahustunud ainete ioone. (Rakumembraanide puhul ei ole mõiste "poolläbilaskvus" kasutamine täiesti õige, kuna see kontseptsioon eeldab, et membraan läbib ainult lahustit molekule, säilitades samal ajal kõik lahustunud ainete molekulid ja ioonid.)
Välimine rakumembraan (plasmalemma) on 7,5 nm paksune ultramikroskoopiline kile, mis koosneb valkudest, fosfolipiididest ja veest. Elastne kile, veest hästi niisutatud ja pärast kahjustusi kiiresti taastav. Sellel on universaalne struktuur, mis on tüüpiline kõikidele bioloogilistele membraanidele. Selle membraani piiripositsioon, osalemine selektiivse läbilaskvuse, pinotsütoosi, fagotsütoosi, eritusproduktide eritumise ja sünteesi protsessides koos naaberrakkudega ja raku kaitsmine kahjustuste eest muudab selle rolli äärmiselt oluliseks. Loomarakud väljaspool membraani on mõnikord kaetud õhukese kihiga, mis koosneb polüsahhariididest ja valkudest, glükokalüksist. Taimerakkudel väljaspool rakumembraani on tugev rakusein, mis loob välise toe ja hoiab raku kuju. See koosneb kiudainetest (tselluloosist), vees lahustumatust polüsahhariidist.