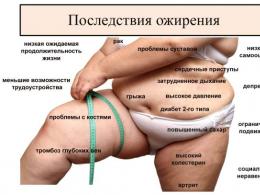
Какие элементы относятся к скелетным мышцам. Строение мышц человека. Скелетная мышечная ткань
Мембранный потенциал поперечно-полосатых мышечных волокон - (-80)- (-90) мВ, а пороговый уровень деполяризации - около -50 мВ ПД, возникая на постсинаптичній мембране мышечного волокна, распространяется сарколемою (мембраной, которая окружает мышечное волокно) в обе стороны от места образования (синапса). Он передается сарколемою електрогенно (аналогично передаче ПД безм"якушевим нервным волокном). Длительность ПД в большинстве скелетных мышц - 2-3 мс. В связи с этим, а также с необходимостью большей поляризации мембраны для возникновения спайка (МП КР = 40 мВ), скорость распространения ПД мембраной мышечного волокна составляет около 3-5 м1с. Через короткое время после поступления ПД мышечное волокно начинает сокращаться. Чтобы понять механизм сокращения мышцы, необходимо ознакомиться с его микроструктурой.
Структура мышечного волокна
Мышечное волокно в диаметре не превышает 0,1 мм, а длина его может составлять от нескольких миллиметров до 12 см (рис. 20).
Под световым микроскопом видно чередование темных и светлых полос (поперечная посмугованість). Темные диски (анизотропные диски - А) имеют двойную променезаломлюваність, светлые (изотропные диски - И) этого свойства не имеют. Часть мышечного волокна от середины одного изотропного диска до середины другого называют саркомером. Длина саркомера в мышце в состоянии покоя - около 2 мкм, а в сокращенном с максимальной силой-немного больше чем 1 мкм. (На рис. 20 изображен саркомер, ограниченный с двух сторон 2-линиями; И - изотропный диск; А - анизотропный диск; Н - участок с уменьшенной анізотропністю. Поперечный срез миофибриллы (д) дает представление о гексагональный распределение толстых и тонких міофіламентів).
Сарколема. Мембрана мышечного волокна - сарколема - образована типичной плазматичною мембраной, укрепленной соединительнотканными волокнами. Последние, сочетаясь у концов мышечных волокон, образуют сухожилки, с помощью которых мышца прикрепляется к костям.
Саркоплазма. В саркоплазмі мышечного волокна находится типичный набор органоидов. Но на особое внимание заслуживает один из них - саркоплазматич-
Рис. 20. в состав мышцы (а) входят мышечные волокна (б), каждое из которых содержит миофибриллы (в). Міофібрила (г) образована из толстых и тонких міофіламентів (г, д)
нийретикулум (СР). Это широко разветвленная сеть, состоящая из цистерн и трубочек, ограниченных двухслойными белково-ліпідними мембранами (рис. 21). Саркоплазматический ретикулум выполняет важную функцию в инициации сокращения мышцы как депо Са2+.

Рис. 21. (по Б.И. Ходоровим): а - распределение трубок (Т-системы) и СР внутри саркомера; б - триада: во время распространения ПД Т-трубкой из цистерны СР выделяются Са2 которые, связываясь с тропоніном в комплексе тропонін-тропоміозин, устраняют тормозное влияние на актиновий міофіламент. Поперечные мостики міозинових филаментов могут теперь взаимодействовать с актиновими філамснтами. Процесс расслабления связан с активным возвращением Са2+ в цистерны
Необходимо упомянуть и о наличии в саркоплазмі белка миоглобина, что служит депо кислорода внутри волокна.
Сократительные протофібрили. Внутри мышечного волокна в саркоплазмі упорядоченно располагаются сократительные протофібрили. Различают протофібрили двух типов: толстые (толщиной 15-17 нм) и тонкие (толщиной около 6 нм). Тонкие протофібрили расположены в И-зоне и с белковыми актиновими нитками. Толстые нити, которые размещены в зоне А, называют міозиновими (см. рис. 20).
Более двухсот молекул миозина участвуют в образовании міозинових филаментов (скрученные попарно, имеют виступний отросток головку). Головки направлены под углом от центра в сторону тонких нитей (напоминают "ерша" для мытья посуды). В основе головки миозина содержится фермент АТФаза, а на самой головке размещается молекула АТФ.
Лктинові філаменти скомпоновано из двух актинових нитей глобулярних молекул актина, имеют вид бусинок. Тонкие нити имеют активные центры, расположены друг от друга на расстоянии 40 нм, к которым могут прикрепляться головки миозина. Кроме актина в тонких нитях содержатся и другие белки - тропониновый комплекс (кальмодулин), который размещается над активными центрами, прикрывая их, что препятствует соединению актина с міозином.
Тонкие нити проходят через середину И-зоны в два близлежащих саркомери. Посередине этой зоны размещена Х-мембрана, что отделяет саркомери друг от друга. Таким образом, содержимое каждого саркомера изолированный сарколемою и Z-мембранами.
Механизм мышечного сокращения
Инициация мышечного сокращения. Распространяясь по наружной мембране, ПД заходит внутрь мышечного волокна (см. рис. 21), здесь он передается на мембрану саркоплазматичного ретикула, где открывает електрозбудливі кальциевые каналы. За то, что в саркоплазмі концентрация кальция меньше чем 10~7 моль1л, а в саркоплазматичному ретикулі - более 10 4 моль1л, начинается интенсивный выход его ионов в саркоплазме.
Выделенный кальций становится инициатором мышечного сокращения. Достаточный для начала мышечного сокращения уровень кальция достигается через 12-15 мс после прихода нервного импульса. Это скрытый, латентный, время мышечного сокращения. В связи с тем, что скорость распространения ПД сарколемою больше времени, необходимого для выделения Са2" из саркоплазматичного ретикула, все фибриллы участка мышцы, иннервируемых одним нервом, сокращаются одновременно.
В инициации мышечного сокращения после поступления в саркоплазме Са2+ определенную роль играет кальмодулин. Присоединяя Са2+, кальмодулин способствует активации Атфазы и использованию энергии АТФ для связи активного центра актинової нити с головкой миозина, а также укорочению мышцы (рис. 22). При соединении кальмодулина (тропонін С) с кальцием активный центр актина высвобождается, вследствие чего к нему присоединяется головка миозина. Эти процессы происходят в том случае, если концентрация свободного кальция в саркоплазмі возрастает в 100 и более раз: с 10"7 до 10~5 моль1л.
"Шарнирный механизм". Вследствие сочетания указанных процессов происходит:
а) подтягивание нитей миозина к атинових;
б) зарядка миозина энергией, которое применяется для выполнения поворота міозинової головки.

Рис. 22. а - поперечные мостики в состоянии расслабления мышечного волокна; 6 - во время сокращения (стрелками указано направление движения актинових протофібрил (и) двух половин саркомера); в - модель развития напряжения в поперечных мостиках
время их сокращения (слева - в состоянии расслабления, справа - во время сокращения мышечного волокна). 4 - шейка поперечного мостика; 5 - головка поперечного мостика
После этого образуемые фосфор и аденозиндифосфорна кислота (АДФ) уходят, а на их место присоединяется новая молекула АТФ, что приводит к разрыву связи миозина с активным центром актина.
При сокращении мышцы:
а) актинові и міозинові нити практически не укорачиваются;
б) взаимодействие актина с міозином приводит к взаимного вхождения нитей в промежутки между ними;
в) две прилегающие 7-мембраны сближаются друг с другом, и при максимально сильном сокращении расстояние между ними может уменьшиться почти вдвое;
г) при уменьшении длины мышцы саркомер расширяется, поскольку заключена внутри саркомера саркоплазма не сжимается;
г) подобные процессы одновременно протекают во всех саркомерах мышечного волокна, поэтому оба конца мышцы подтягиваются к центру.
в настоящее время еще окончательно неизвестен механизм, который обеспечивает вхождение актоміозинових нитей друг в друга. Общепринятая гипотеза "шарнирного механизма" (см. рис. 22). После соединения головки миозина с активным центром актина происходит ее поворот на 45°. Вследствие разрыва мостика шейка головки миозина выпрямляется, приобретая исходное положение. За такие движения эта система и получила название шарнирного механизма. Во время поворота миозин продвигается актином на один "шаг", или "гребок", равен 20 нм. Поступления новой порции Са2+ приводит к повторению "шага", но теперь уже другой головки, что оказалась напротив нового активного центра актина, поскольку они расположены на расстоянии около 40 нм друг от друга. В связи с тем что міозинові нити имеют биполярную организацию головок, то параллельные "гребки" их обеспечивают скольжение актинових нитей вдоль саркомера (от мембраны до его середины).
Расслабление мышцы.
Указанные процессы ("шаги") будут повторяться до тех пор, пока саркоплазма содержит свободный Са2" (в концентрации более 10-5 моль1л) и АТФ. Если нет новой волны деполяризации, кальций быстро возвращается обратно в цистерны саркоплазматичного ретикула. Он откачивается из саркоплазми против градиента концентрации с помощью Са2+-насоса, расположенного на мембране саркоплазматичного ретикула. Работа этого насоса, что требует большого количества АТФ (для удаления каждого Са2+ используется 2 молекулы АТФ), активируется самим кальцием, точнее, ростом его концентрации в саркоплазмі. Следствие откачивания кальция из саркоплазми - разрыв всех связей актина и миозина и расслаблению мышцы.
Энергетика мышечного сокращения
АТФ в мышце необходима для:
1) сокращения (образования мостиков);
2) расслабления (разрыва мостиков);
3) работы Са2+-насоса;
4) работы К* - насоса (для устранения нарушенных ионных градиентов вследствие поступления возбуждение).
Однако в саркоплазмі мышцы АТФ относительно немного. ее хватит лишь на несколько мышечных сокращений (примерно восемь одиночных сокращений). Вместе с тем в естественных условиях мышцы могут сокращаться длительное время, что становится возможным лишь благодаря активации механизмов ре-синтеза АТФ креатинфосфокіназного, гліколітичного, аэробного окисления.
Последовательность "включения" указанных путей ресинтеза АТФ такая. Сначала, сразу после гидролиза АТФ, начинается ее восстановление за счет креатинфосфата (КФ):
АДФ + КФ <=>АТФ + КФ.
Креатинфосфокіназний путь безынерционный (он запускается сразу АДФ, что образуется) и может обеспечить сокращение мышц в течение нескольких секунд. Одновременно с ним активируется гліколітичний путь. Образование АТФ во время гликолиза углеводов происходит при участии ферментов, активность которых возрастает постепенно от начала мышечных сокращений. Но уже через 15-20 с они становятся достаточно активными, чтобы подхватить эстафету ресинтеза АТФ при истощении КФ. Недостаток этого пути - меньший выход АТФ за единицу времени по сравнению с предыдущим. Кроме того, во время гликолиза образуются недоокиснені продукты (молочная, пировиноградная кислоты), что в случае интенсивного образования не успевают выйти из мышцы, приводя к нарушению в нем гомеостаза (сдвиг рН в кислую сторону).
Наибольшие возможности для ресинтеза АТФ имеет аэробное окисление (практически неограниченное время при адекватном поступлении кислорода и продуктов окисления). Но это найінерційніший путь, так как его ферментная система активируется медленно. На максимальный уровень активности она выходит через 2-3 мин от начала мышечной работы. Кроме митохондриальных ферментов самого мышечного волокна обеспечения указанного механизма ресинтеза АТФ требует адекватного снабжения мышц кислородом и исходными продуктами. К тому же производительность (количество синтезированной АТФ за единицу времени) аэробного окисления неодинакова в зависимости от соединения, окисляется: при окиснюванні углеводов энергии.
Естественно, что указанные возможности путей ресинтеза АТФ определяют работоспособность мышц.
Коэффициент полезного действия и теплообразование при мышечной работе
Согласно первому закону термодинамики (закон сохранения энергии) химическая энергия превращается в мышце, равна сумме механической энергии (работы) и теплотворность. Гидролиз одного моля АТФ обеспечивает получение 48 кДж энергии. Лишь 40-45 % ее превращается в механическую энергию, а остальные 55-60 % превращаются в на первоначальное тепло. Однако в природных условиях механическая эффективность мышечной деятельности, или коэффициент полезного действия, не превышает 20-30 %. Это обусловлено тем, что не вся энергия АТФ в мышце идет на собственно мышечное сокращение: часть ее расходуется на процессы восстановления. Следовательно, чем выше интенсивность мышечной работы, то более активные процессы теплообразование.
Типы и режимы мышечных сокращений
В естественных условиях оба конца мышцы прикрепляются с помощью сухожилий к костям и при сокращении притягивают их друг к другу. Если один конец мышцы (сустав) закреплен, то к нему подтягивается другой (рис. 23). Когда на этом конце мышцы прикреплен груз, который мышца поднять не в СОСТОЯНИИ, то он только напрягается, в таком случае его длина не меняется. Случаются и такие состояния, когда мышца постепенно увеличивается в длину (груз тяжелее, чем подъемная сила мышцы, или необходимо медленно опустить груз).
В экспериментальных условиях можно выделить один мускул, одно волокно и даже одну актоміозинову нить с нервом, иннервирует, или без него. Если закрепить один конец в штативе неподвижно, а к другому подвесить груз или устройство для регистрации, можно записать сокращение мышцы - міограму.
Вследствие этого различают следующие типы мышечных сокращений:
o ізотонічне (концентрическое) - сокращение мышц с укорочением при сохранении постоянного напряжения;
o изометрическое, когда длина мышцы не изменяется (напряжение);
o эксцентричное (плиометрическая), когда мышца удлиняется.
Как правило, большинство естественных сокращений мышц смешанные, то есть анізотонічного типа, когда мышца укорачивается в случае повышения напряжения.
На рис. 24, а приведены кривую одиночного сокращения. На ней можно различить фазы сокращения и расслабления. Вторая фаза более длительная. Время одного сокращения даже одиночного волокна значительно превышает время существования ПД.

Рис. 24. Разнообразные режимы сокращения мышц:
а - одиночные сокращения; в - неполный тетанус; г г - полный тетанус

Рис. 23. Взаимодействие мышц-сгибателей (а) и мышц-разгибателей (б)
Амплитуда одиночного сокращения изолированного мышечного волокна не зависит от силы раздражения, а подчиняется закону "все или ничего". В отличие от этого на сплошном мышце можно получить "лестница" (лестница Боудича): что большей силы (до определенной величины) раздражение, тем сильнее сокращение. Дальнейшее увеличение силы раздражения не влияет на амплитуду сокращения мышцы. Указанную закономерность прослеживают как при раздражении через нерв, так и в случае раздражения самой мышцы. Это обусловлено тем, что практически все мышцы (и нервы) смешанные, то есть состоят из множества двигательных единиц (РО), имеющих различную возбудимость.
Двигательная единица
Одиночное нервное волокно мотонейрона и мышечные волокна, что им иннервируются, составляют одну двигательную единицу (рис. 25). В большинстве скелетных мышц к двигательной единицы входит несколько сотен (даже тысяч) мышечных волокон. Даже в очень маленьких мышцах, от которых требуют высокой точности движений (глаз, кисти), в составе двигательной единицы может насчитываться 10-20 мышечных волокон. С функциональной точки зрения различают несколько типов двигательной единицы, которые можно сгруппировать следующим образом: быстрые и медленные. Функциональные их различия обусловлены соответствующими структурными особенностями, к тому же как на уровне сравнительно грубой морфологии, так и тонкого биохимического дифференцирования. Различные типы двигательных единиц отличают мышечные части, так и нервные волокна. Эти отличия и обеспечивают соответствующее функциональное проявление каждого типа двигательных единиц. Быстрые и медленные отличаются возбудимостью, скоростью проведения импульсов аксоном, оптимальной частотой импульсации и устойчивостью к утомлению после выполнения работы. К тому же в каждом типе мотонейрон и мышечные волокна как партнеры соединены друг с другом, что и обеспечивает их функциональные особенности.
Мотонейроны. Возбудимость или чувствительность к силе действующего раздражителя, мотонейронов одного и того же мышцы находится в обратной зависимости

Рис. 25.
1 - тело мотонейрона; 2 - ядро; С - дендриты; 4 - аксон; 5 - миелиновая оболочка аксона; 6 - концевые веточки аксона; 7 - нервно-мышечные синапсы
от размеров их тела: чем меньше мотонейрон, тем выше его возбудимость, то есть при меньшей силе раздражителя в них появляется ПД. Маленькие мотонейроны иннервируют сравнительно незначительное количество медленных мышечных волокон, большие - быстрые мышечные волокна, которых в одной двигательной единицы, как правило, много.
От величины нейрона зависит диаметр аксона и скорость проведения возбуждения по нему: она выше в больших мотонейронах. Кроме того, в таких мотонейронах могут возникать нервные импульсы с большой частотой. Следовательно, за счет изменения частоты импульсации мотонейрона мышечные волокна, входящие в состав соответствующей двигательной единицы, могут получать высокий частотный диапазон ПД, а это обусловит и большую силу их сокращения.
Каждому мотонейрону соответствует и структура мышечных волокон двигательной единицы. Так, скорость сокращения мышечного волокна находится в прямой зависимости от активности актоміозинової Атфазы (количества актинових и міозинових филаментов): чем выше ее активность, тем быстрее образуются актоміозинові мостики, и следовательно, выше скорость сокращения. Плотность "упаковка" актоміози новых филаментов в быстрых мышечных волокнах выше, чем в медленных. Кроме того, в скором волокне более выражен саркоплазматический ретикулум (депо кальция). Поэтому во время поступления ПД:
o скрытый время до начала сокращения меньший;
o плотность кальциевого насоса выше.
Итак, мышца быстрее сокращается и расслабляется. В быстром мышечном волокне повышена активность ферментов гликолиза, которые обеспечивают быстрое восстановление АТФ, которая расходуется во время выполнения интенсивных мышечных сокращений.
В отличие от этого в медленном мышечном волокне выше активность ферментов окисления, благодаря которым восстановление АТФ происходит хоть и медленнее, но зато экономически выгоднее. Так, если из 1 моля глюкозы в результате гликолиза образуется лишь 2-3 моля АТФ, то в случае аэробного окисления - 36-38 моль АТФ. Кроме того, во время гликолиза образуются недоокиснені субстраты (например, молочной кислоты), что "закиснюють" мышцу и снижают его работоспособность. Повышению работоспособности, улучшению условий окисления способствуют еще две структурные различия медленных мышечных волокон:
1) медленные волокна лучше, чем быстрые, обеспечиваются кислородом благодаря большей плотности кровеносных капилляров, окружающих их;
2) внутри медленных волокон содержится большое количество миоглобина, который придает им красную окраску и являются депо кислорода, что может применяться для окисления в момент мышечного сокращения, когда поступление кислорода с кровью затрудняется из-за сжатия кровеносных сосудов мышцей, которая сокращается.
Быстрые мышечные волокна имеют короткий период сокращения - до 7,5 мс, а медленные - довгий - до 100 мс.
Таким образом, подытоживая функциональные различия двигательных единиц, можно отметить: медленные двигательные единицы отличаются легкой возбудимостью, меньшей силой и скоростью сокращения при малой утомляемости и высокой выносливости. Быстрые двигательные единицы имеют противоположные свойства.
Исследования последних лет доказали, что каждый человек имеет врожденные различия процентного соотношения быстрых и медленных волокон в скелетных мышцах. Например, во внешнем мышце бедра диапазон колебания количества медленных волокон составляет от 13 до 96 %. Преимущество медленных волокон обеспечивает "стайерские", а малый их процент - "спринтерские" возможности спортсмена. К тому же компоновка различных мышц одного человека также различается. Так, в среднем содержание медленных волокон в триголовому мышце плеча составляет 33 %, двуглавому - 49, переднем великогомілковому - 46, камбало-образном - 84 %.
Суммация сокращения и тетанус
В естественных условиях жизни человека одиночных сокращений мышц не бывает. Обычно к мышцам нервные импульсы по мотонейронах поступают "пачками", то есть подряд по несколько штук с относительно незначительными временными промежутками. Это приводит к формированию не одного, а нескольких ПД и в самом скелетном мышце. Если на мышцу действуют не одиночные импульсы (ПД), а такие, которые быстро идут один за одним, то сократительные эффекты суммируются, и в результате этого мышца сокращается длительное время (см. рис. 24). К тому же если дальнейшие раздражители поступают в начальный момент расслабления, міографічна кривая будет зубчатой, а если до начала расслабления-без зазубрин. Такой тип сокращений называют тетанусом.
Различают зубчатый и непосмугований тетануси. Во время тетанусу не только удлиняется время сокращения, но и растет его сила. Это происходит из-за того, что в ответ на первый ПД успеют произойти лишь незначительные "шаги". Окончательный резерв создает возможность увеличить силу сокращения во время поступления дальнейших ПД. В этом случае концентрация кальция (количество актоміозинових мостиков) в таком мышечном волокне может быть такой же, как и во время одиночного сокращения.
Тетанічне сокращение вероятно прежде всего потому, что мембрана мышечного волокна способна проводить довольно частые ПД (более 100 в 1 с), поскольку рефрактерный период в скелетных мышцах гораздо короче собственно одиночного сокращения. Следовательно, когда к мышце поступают следующие ПД, он снова становится чувствителен к ним.
Частота и сила раздражителя, необходимые для выведения мышечного волокна в тетанус, не для всех мышц одинаковы, а зависят от особенностей их двигательной единицы. Продолжительность одного сокращения медленного мышечного волокна может достигать 100 мс, а быстрого - 10-30 мс. Поэтому для получения непосмугованого тетанусу в медленных волокнах достаточно 10-15 имп.1с, а быстрые нуждаются до 50 имп.1с и выше.
В природных условиях почти никогда не случается, чтобы все мышечные волокна находились в сокращенном состоянии. Поэтому при произвольном сокращении сила мышц меньше, чем в случае искусственного раздражения. На этом принципе основан механизм резкого повышения силы сокращения мышц в экстремальной ситуации: возрастает синхронность нервных импульсов, поступающих к различной двигательной единицы. Один из механизмов, обеспечивающих увеличение мышечной силы, например спортсмена во время тренировки, - рост синхронности сокращения отдельных двигательных единиц.
Максимальный ритм возбуждения. Предельный ритм возбуждения, обусловленный понятием лабильность, всех возбудимых тканей зависит от продолжительности периода, необходимого для восстановления чувствительности натриевых каналов после предыдущего раздражения, то есть от периода рефрактерности. Лабильность двигательной единицы, состоящий из трех структур (нерва, синапса, мышцы), определяют наиболее "узкой" звеном-синапсом, поскольку именно он имеет минимальную частоту передачи возбуждения. Мотонейроны, даже самые маленькие, способны проводить более 200 имп.1с, мышечные волокна-более 100 имп.1с, а нервно-мышечный синапс - меньше чем 100 имп.1с.
Функциональная характеристика скелетных мышц
Силу мышцы определяют силой тяги на ее концах. Максимальная сила тяги развивается во время изометрического сокращения мышцы при соблюдении следующих условий: а) активации всех двигательных единиц, из которых состоит эта мышца; б) начала сокращения мышцы при длине покоя; в) режима полного тетанусу во всех двигательных единиц.

Рис. 26. (по А.А. Ухтомским)
Для измерения силы мышцы определяют либо максимальный груз, который он сможет поднять, либо максимальное напряжение, которое он сможет развить в условиях изометрического сокращения. (Одиночное мышечное волокно способно развить напряжение 100-200 мг.) В теле человека содержится около 30 млн мышечных волокон и теоретически, если бы все они тянули в одну сторону, то создали бы напряжение до 30 т. Кроме того, необходимо учитывать еще и такие обстоятельства. Во-первых, сила различных мышечных волокон несколько отличается: быстрые двигательные единицы сильнее медленных. Во-вторых, сила мышцы зависит от ее поперечного сечения: чем больше объем мышцы, то он сильнее. К тому же в зависимости от хода волокон различают мышцы косые и прямые. Косой ход волокон обеспечивает большое количество мышечных волокон, проходящих через ее поперечное сечение, вследствие чего сила такого мышцы больше. Поэтому различают физиологический и анатомический поперечника мышцы: физиологический поперечник перпендикулярен к направлению мышечных волокон, а анатомический - до длины мышцы (рис. 26). Естественно, что в мышц с продольным направлением волокон оба названных поперечника совпадают, а в перистых - физиологический поперечник больше, чем анатомический, поэтому за одинакового анатомического поперечника последние сильнее. Например, относительная сила мышц человека (на 1 см2 площади поперечного сечения):
o голеностопный мышца - 5,9 кг;
o мышцу - сгибатель плеча - 8,1 кг;
o жевательный мускул - 10,0 кг;
o двуглавая мышца плеча - 11,4 кг;
o трехглавую мышцу плеча - 16,7 кг.
В естественных условиях на проявление мышечной силы влияют не только указанные выше три условия, но и угол, под которым мышца подходит к кости. Что больший угол прикрепления, то лучшие условия для проявления силы. Если мышца подходит к кости под прямым углом, почти вся его сила расходуется на обеспечение движения, а если под острым - часть силы идет на обеспечение движения, остальные - на сжатие рычага.
Утомление
Во время длительной или интенсивной мышечной работе развивается утомление, которое выражается вначале в снижении работоспособности, а затем и в прекращении работы. Утомление характеризуется соответствующими изменениями, которые возникают не только в мышцах, но и в системах, обслуживающих их.
Утомлением называют состояние, развивающееся вследствие работы и проявляется ухудшением двигательных и вегетативных функций организма, их координации. В этом случае снижается работоспособность, появляется чувство устали (психологическое состояние). Утомление - целостная реакция всего организма. Поэтому, когда ниже будет рассматриваться утомление нерва, синапса, мышц, необходимо помнить об условности этих понятий. Правильнее говорить о некоторых механизмах, определяющих "работоспособность" основных звеньев двигательной единицы - нервных, мышечных волокон, синапса.
Утомление нервного волокна. В природных условиях нервное волокно практически не устает. Проведение нервного импульса требует затраты энергии только для работы К+-насоса, что достаточно енергоекономічно. Системы ресинтеза АТФ вполне справляются с обеспечением энергией нервного волокна.
Утомление нервно-мышечного синапса. Работоспособность, то есть способность проводить возбуждение, у синапса значительно ниже, чем у нервного волокна. Это может быть следствием двух явлений. Депрессия передачи возбуждения в синапсе может спричинюватися истощением значительной части медиатора или ослаблением его восстановления при слишком высокой частоте ПД, поступающих нервным волокном. Кроме того, при интенсивной мышечной деятельности недоокиснені продукты (активно образуются во время гликолиза) снижают чувствительность постсинаптичної мембраны к медиатору АХ. Это приводит к снижению амплитуды каждого ПКП и при чрезмерном снижении возникновения ПД вообще становится невозможным.
Утомление мышечного волокна. Нарушение возбудимости и сократимости мышечного волокна в первую очередь обусловлено нарушением его энергетики, то есть механизмов ресинтеза АТФ. В этом случае решающим моментом становится интенсивность мышечной работы. Сверхвысокая ее активность связана с дефицитом креатинфосфокіназного пути или накоплением недоокисленных продуктов во время гликолиза. Последнее, с одной стороны, снижает чувствительность постсинаптичної мембраны, с другой - сдвигает рН саркоплазми в кислую сторону, что само по себе тормозит активность гликолитических ферментов. Все это и вызывает быстрое развитие утомления при интенсивной мышечной работе. Утомление во время длительной малоінтенсивної работы развивается медленно, что связано с нарушением механизмов регуляции со стороны центральных отделов нервной системы.
Скелетные мышцы построены из поперечнополосатой скелетной мышечной ткани. Они являются произвольными, т.е. их сокращение осуществляется сознательно и зависит от нашего желания. Всего в теле человека насчитывается 639 мышц, 317 из них - парные, 5 - непарные.
Скелетная мышца - это орган, имеющий характерную форму и строение, типичную архитектонику сосудов и нервов, построенный в основном из поперечнополосатой мышечной ткани, покрытый снаружи собственной фасцией, обладающий способностью к сокращению.
Принципы классификации мышц . В основу классификации скелетных мышц человеческого организма положены различные признаки: область тела, происхождение и форма мышц, функция, ана-
томо-топографические взаимоотношения, направление мышечных волокон, отношение мышцы к суставам. По отношению к областям человеческого тела различают мышцы туловища, головы, шеи и конечностей. Мышцы туловища в свою очередь разделяют на мышцы спины, груди и живота. Мышцы
верхней конечности соответственно имеющимся частям скелета делят на мышцы пояса верхней конечности, мышцы плеча, предплечья и кисти. Гомологичные отделы характерны для мышц нижней конечности - мышцы пояса нижней конечности (мышцы таза), мышцы бедра, голени и стопы.
По форме мышцы могут быть простыми и сложными. К простым мышцам относят длинные, короткие и широкие. Сложными считают многоглавые (двуглавые, трехглавые, четырехглавые), многосухожильные, двубрюшные мышцы. Сложными являются также мышцы определенной геометрической формы: круглые, квадратные, дельтовидные, трапециевидные, ромбовидные и т. д.
По функции различают мышцы-сгибатели и разгибатели; мышцы приводящие и отводящие; вращающие (ротаторы); сфинктеры (суживатели) и дилятаторы (расширители). Вращающие мышцы в
зависимости от направления движения подразделяют на пронаторы и супинаторы (вращающие внутрь и наружу). Также предусматривается подразделение их на синергисты и антагонисты. Синергисты - это мышцы, выполняющие одинаковую функцию и при этом усиливающие друг друга. Антагонисты - это мышцы, выполняющие противоположные функции, т.е. производящие противоположные друг другу движения.
По расположению - поверхностные и глубокие; наружные и внутренние; медиальные и латеральные.
По направлению мышечных волокон - с параллельным, косым, круговым и поперечным ходом мышечных волокон.
Строение мышц. Скелетная мышца как орган включает в себя собственно мышечную и сухожильную части, систему соединительнотканных оболочек, собственные сосуды и нервы. Средняя, утолщенная часть мышцы называется брюшком. На обоих концах мышцы в большинстве случаев находятся сухожилия, с помощью которых она прикрепляется к костям. Структурно-функциональной единицей собственно мышечной части является поперечнополосатое мышечное волокно .
В процессе мышечного сокращения актиновые нити втягиваются в промежутки между миозиновыми, изменяют свою конфигурацию, сцепляются друг с другом. Обеспечение энергией этих процессов происходит за счет расщепления в митохондриях молекул АТФ.
Функциональная единица мышцы - мион - совокупность поперечнополосатых мышечных волокон, иннервируемых одним двигательным нервным волокном. Вспомогательным аппаратом скелетных мышц являются фасции, фиброзные и костно-фиброзные каналы, синовиальные влагалища, синовиальные сумки, мышечные блоки и сесамовидные кости. Фасции представляют собой соединительнотканные оболочки, ограничивающие подкожную жировую клетчатку, покрывающие мышцы и некоторые внутренние органы.
Структурно-функциональной единицей скелетной мышцы является симпласт или мышечное волокно - огромная клетка, имеющая форму протяженного цилиндра с заостренными краями (под наименованием симпласт, мышечное волокно, мышечная клетка следует понимать один и тот же объект).
Длина мышечной клетки чаще всего соответствует длине целой мышцы и достигает 14 см, а диаметр равен нескольким сотым долям миллиметра.
Мышечное волокно , как и любая клетка, окружено оболочкой - сарколемой. Снаружи отдельные мышечные волокна окружены рыхлой соединительной тканью, которая содержит кровеносные и лимфатические сосуды, а так же нервные волокна.
Группы мышечных волокон, образуют пучки, которые, в свою очередь, объединяются в целую мышцу, помещенную в плотный чехол соединительной ткани переходящей на концах мышцы в сухожилия, крепящиеся к кости (рис.1).

Рис. 1.
Усилие, вызываемое сокращением длины мышечного волокна, передается через сухожилия костям скелета и приводит их в движение.
Управление сократительной активностью мышцы осуществляется с помощью большого числа мотонейронов (рис. 2) - нервных клеток, тела которых лежат в спинном мозге, а длинные ответвления - аксоны в составе двигательного нерва подходят к мышце. Войдя в мышцу, аксон разветвляется на множество веточек, каждая из которых подведена к отдельному волокну.

Рис. 2.
Таким образом, один мотонейрон иннервирует целую группу волокон (так называемая нейромоторная единица), которая работает как единое целое.
Мышца состоит из множества нервно моторных единиц и способна работать не всей своей массой, а частями, что позволяет регулировать силу и скорость сокращения.
Для понимания механизма сокращения мышцы необходимо рассмотреть внутреннее строение мышечного волокна, которое, как вы уже поняли, сильно отличается от обычной клетки. Начнем с того, что мышечное волокно многоядерно. Связано это с особенностями формирования волокна при развитии плода. Симпласты (мышечные волокна) образуются на этапе эмбрионального развития организма из клеток предшественников - миобластов.
Миобласты (неоформленные мышечные клетки) интенсивно делятся, сливаются и образуют мышечные трубочки с центральным расположением ядер. Затем в мышечных трубочках начинается синтез миофибрилл (сократительных структур клетки см. ниже), и завершается формирование волокна миграцией ядер на периферию. Ядра мышечного волокна к этому времени уже теряют способность к делению, и за ними остается только функция генерации информации для синтеза белка.
Но не все миобласты идут по пути слияния, часть из них обособляется в виде клеток-сателлитов, располагающихся на поверхности мышечного волокна, а именно в сарколеме, между плазмолемой и базальной мембраной - составными частями сарколемы. Клетки-сателлиты, в отличие от мышечных волокон, не утрачивают способность к делению на протяжении всей жизни, что обеспечивает увеличение мышечной массы волокон и их обновление. Восстановление мышечных волокон при повреждении мышцы возможно благодаря клеткам-сателлитам. При гибели волокна, скрывающиеся в его оболочке, клетки-сателиты активизируются, делятся и преобразуются в миобласты.
Миобласты сливаются друг с другом и образуют новые мышечные волокна, в которых затем начинается сборка миофибрилл. То есть при регенерации полностью повторяются события эмбрионального (внутриутробного) развития мышцы.
Помимо многоядерности отличительной чертой мышечного волокна является наличие в цитоплазме (в мышечном волокне ее принято называть саркоплазмой) тонких волоконец – миофибрилл (рис.1), расположенных вдоль клетки и уложенных параллельно друг другу. Число миофибрилл в волокне достигает двух тысяч.
Миофибриллы являются сократительными элементами клетки и обладают способностью уменьшать свою длину при поступлении нервного импульса, стягивая тем самым мышечное волокно. Под микроскопом видно, что миофибрилла имеет поперечную исчерченность - чередующиеся темные и светлые полосы.
При сокращении миофибриллы светлые участки уменьшают свою длину и при полном сокращении исчезают вовсе. Для объяснения механизма сокращения миофибриллы около пятидесяти лет назад Хью Хаксли была разработана модель скользящих нитей, затем она нашла подтверждение в экспериментах и сейчас является общепринятой.
ЛИТЕРАТУРА
- МакРоберт С. Руки титана. – М.: СП " Уайдер спорт", 1999.
- Остапенко Л. Перетренированность. Причины возникновения перетренированности при силовом тренинге // Ironman, 2000, № 10-11.
- Солодков А. С., Сологуб Е. Б. Физиология спорта: Учебное пособие. – СПб: СПбГАФК им. П.Ф. Лесгафта, 1999.
- Физиология мышечной деятельности: Учебник для институтов физической культуры / Под ред. Коца Я. М. – М.: Физкультура и спорт, 1982.
- Физиология человека (Учебник для институтов физической культуры. Изд. 5-е). / Под ред. Н. В. Зимкина. – М.: Физкультура и спорт, 1975.
- Физиология человека: Учебник для студентов медицинских институтов / Под ред. Косицкого Г. И. - М.: Медицина, 1985.
- Физиологические основы спортивной тренировки: Методические указания по спортивной физиологии. – Л.: ГДОИФК им. П.Ф. Лесгафта, 1986.
КЛАССИФИКАЦИЯ МЫШЕЧНЫХ ВОЛОКОН.
Морфологическая классификация
Поперечно-полосатая (поперечно-исчерченная)
Гладкая (неисчерченная)
Классификация по типу контроля мышечной актичности
Поперечно-полосатая мышечная ткань скелетного типа.
Гладкая мышечная ткань внутренних органов.
Поперечно-полосатая мышечная ткань сердечного типа
КЛАССИФИКАЦИЯ СКЕЛЕТНЫХ МЫШЕЧНЫХ ВОЛОКОН
ПОПЕРЕЧНО-ПОЛОСАТЫЕ МЫШЦЫ представляют собой максимально специализированый аппарат для осуществления быстрого сокращения. Поперечно-полосатые мышцы бывают двух типов - скелетные и сердечные. СКЕЛЕТНЫЕ мышцы состоят из мышечных волокон, каждое из которых представляет собой многоядерную клетку, полученную в результате слияния большого количества клеток. В зависимости от сократительных свойств, окраски и утомляемости мышечные волокна подразделяют на две группы - КРАСНЫЕ И БЕЛЫЕ. Функциональной единицей мышечного волокна является миофибрилла. Миофибриллы занимают практически всю цитоплазму мышечного волокна, оттесняя ядра на периферию.
КРАСНЫЕ МЫШЕЧНЫЕ волокна (волокна 1 типа) содержат большое количество митохондрий с высокой активностью окислительных ферментов. Сила их сокращений сравнительно невелика, а скорость потребления энергии такова, что им вполне хватает аэробного метаболизма (используют кислород). Они участвуют в движениях, не требующих значительных усилий, - например, в поддержании позы.
БЕЛЫМ МЫШЕЧНЫМ ВОЛОКНАМ (волокнам 2 типа) присуща высокая активность ферментов гликолиза, значительная сила сокращения и такая высокая скорость потребления энергии, для которой уже не хватает аэробного метаболизма. Поэтому двигательные единицы, состоящие из белых волокон, обеспечивают быстрые, но кратковременные движения, требующие рывковых усилий.
КЛАССИФИКАЦИЯ ГЛАДКИХ МЫШЦ
Гладкие мышцы подразделяются на ВИСЦЕРАЛЬНЫЕ (УНИТАРНЫЕ) И МУЛЬТИУНИТАРНЫЕ . ВИСЦЕРАЛЬНЫЕ ГЛАДКИЕ мышцы находятся во всех внутренних органах, протоках пищеварительных желез, кровеносных и лимфатических сосудах, коже. К МУЛЫПИУНИТАРНЫМ относятся ресничная мышца и мышца радужки глаза. Деление гладких мышц на висцеральные и мультиунитарные основано на различной плотности их двигательной иннервации. В ВИСЦЕРАЛЬНЫХ ГЛАДКИХ мышцах двигательные нервные окончания имеются на небольшом количестве гладких мышечных клеток.
ФУНКЦИИ СКЕЛЕТНЫХ И ГЛАДКИХ МЫШЦ.
ФУНКЦИИ И СВОЙСТВА ГЛАДКИХ МЫШЦ
1. ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ . Гладкие мышцы характеризуются нестабильным мембранным потенциалом. Колебания мембранного потенциала независимо от нервных влияний вызывают нерегулярные сокращения, которые поддерживают мышцу в состоянии постоянного частичного сокращения - тонуса. Мембранный потенциал гладкомышечных клеток не является отражением истинной величины потенциала покоя. При уменьшении мембранного потенциала мышца сокращается, при увеличении - расслабляется.
2. АВТОМАТИЯ . ПД гладких мышечных клеток имеют авторитмический характер, подобно потенциалам проводящей системы сердца. Это свидетельствует о том, что любые клетки гладких мышц способны к самопроизвольной автоматической активности. Автоматия гладких мышц, т.е. способность к автоматической (спонтанной) деятельности, присуща многим внутренним органам и сосудам.
3. РЕАКЦИЯ НА РАСТЯЖЕНИЕ . В ответ на растяжение гладкая мышца сокращается. Это вызвано тем, что растяжение уменьшает мембранный потенциал клеток, увеличивает частоту ПД и в конечном итоге - тонус гладкой мускулатуры. В организме человека это свойство гладкой мускулатуры служит одним из способов регуляции двигательной деятельности внутренних органов. Например, при наполнении желудка происходит растяжение его стенки. Увеличение тонуса стенки желудка в ответ на его растяжение способствует сохранению объема органа и лучшему контакту его стенок с поступившей пищей. В кровеносных сосудах растяжение, создаваемое колебаниями кровяного давления.
4. ПЛАСТИЧНОСТ Ь. Изменчивость напряжения без закономерной связи с ее длиной. Так, если растянуть гладкую мышцу, то ее напряжение будет увеличиваться, однако если мышцу удерживать в состоянии удлинения, вызванным растяжением, то напряжение будет постепенно уменьшаться, иногда не только до уровня, существовавшего до растяжения, но и ниже этого уровня.
5. ХИМИЧЕСКАЯ ЧУВСТВИТЕЛЬНОСТЬ . Гладкие мышцы обладают высокой чувствительностью к различным физиологически активным веществам: адреналину, норадреналину. Это обусловлено наличием специфических рецепторов мембраны гладкомышечных клеток. Если добавить адреналин или норадреналин к препарату гладкой мышцы кишечника, то увеличивается мембранный потенциал, уменьшается частота ПД и мышца расслабляется, т. е. наблюдается тот же эффект, что и при возбуждении симпатических нервов.
ФУНКЦИИ И СВОЙСТВА СКЕЛЕТНЫХ МЫШЦ
Скелетная мускулатура является составной частью опорно-двигательного аппарата человека. При этом мышцы выполняют следующие функции :
1)обеспечивают определенную позу тела человека;
2)перемещают тело в пространстве;
3) перемещают отдельные части тела относительно друг друга;
4) являются источником тепла, выполняя терморегуляционную функцию.
Скелетная мышца обладает следующими важнейшими СВОЙСТВАМИ :
1)ВОЗБУДИМОСТЬЮ - способностью отвечать на действие раздражителя изменением ионной проводимости и мембранного потенциала.
2) ПРОВОДИМОСТЬЮ - способностью проводить потенциал действия вдоль и в глубь мышечного волокна по Т-системе;
3) СОКРАТИМОСТЬЮ - способностью укорачиваться или развивать напряжение при возбуждении;
4) ЭЛАСТИЧНОСТЬЮ - способностью развивать напряжение при растягивании.
Строение мышцы:
А - внешний вид двуперистой мышцы; Б - схема продольного разреза многоперистой мышцы; В - поперечный разрез мышцы; Г - схема строения мышцы как органа; 1, 1" - сухожилие мышц; 2 - анатомический поперечник мышечного брюшка; 3 - ворота мышцы с сосудисто-нервным пучком (а - артерия, в - вена, п - нерв); 4 - физиологический поперечник (суммарный); 5 - подсухожильная бурса; 6-6" - кости; 7 - наружный перимизий; 8 - внутренний перимизий; 9 - эндомизий; 9"-мышечные волокна; 10, 10", 10" - чувствительные нервные волокна (несут импульс от мышцы, сухожилий, сосудов); 11, 11" - двигательные нервные волокна (несут импульс в мышцы, сосуды)
СТРОЕНИЕ СКЕЛЕТНОЙ МЫШЦЫ КАК ОРГАНА
Скелетные мышцы - musculus skeleti - являются активными органами аппарата движения. В зависимости от функциональных потребностей организма они могут изменять взаимоотношения между костными рычагами (динамическая функция) или укреплять их в определенном положении (статическая функция). Скелетные мышцы, выполняя сократительную функцию, значительную часть химической энергии, полученную с пищей, трансформируют в тепловую энергию (до 70%) и в меньшей степени в механическую работу (около 30%). Поэтому при сокращении мышца выполняет не только механическую работу, но и служит основным источником тепла в организме. Вместе с сердечно–сосудистой системой скелетные мышцы активно участвуют в обменных процессах и использовании энергетических ресурсов организма. Наличие в мышцах большого числа рецепторов способствует восприятию мышечно–суставного чувства, которое совместно с органами равновесия и органами зрения обеспечивает выполнение точных мышечных движений. Скелетные мышцы в совокупности с подкожной клетчаткой содержат до 58% воды, выполняя тем самым важную роль основных депо воды в организме.
Скелетная (соматическая) мускулатура представлена большим количеством мышц. Каждая мышца имеет опорную часть - соединительнотканную строму и рабочую часть - мышечную паренхиму. Чем большую статическую нагрузку выполняет мышца, тем больше развита в ней строма.
Снаружи мускул одет соединительнотканной оболочкой, которая называется наружным перимизием
Perimysium . На различных мышцах он разной толщины. От наружного перимизия внутрь отходят соединительнотканные перегородки - внутренний перимизий, окружающий мышечные пучки различной величины. Чем большую статическую функцию несет мышца, тем более мощные соединительнотканные перегородки в ней расположены, тем их больше. На внутренних перегородках в мышцах могут закрепляться мышечные волокна, проходят сосуды и нервы. Между мышечными волокнами проходят очень нежные и тонкие соединительнотканные прослойки, называемые эндомизием -endomysium .
В строме мышцы, представленной наружным и внутренним перимизием и эндомизием, упакована мышечная ткань (мышечные волокна, образующие мышечные пучки), формирующая различной формы и величины мышечное брюшко. Строма мышцы по концам мышечного брюшка образует сплошные сухожилия, форма которых зависит от формы мышц. Если сухожилие шнурообразно, оно называется просто сухожилием - tendo . Если сухожилие плоское, идет от плоского мускульного брюшка, то оно называется, апоневрозом –aponeurosis .
В сухожилии также различают наружные и внутренние оболочки (мезотендиний - mesotendineum ). Сухожилия очень плотны, компактны, образуют прочные шнуры, обладающие большой сопротивляемостью на разрыв. Коллагеновые волокна и пучки в них расположены строго продольно, благодаря чему сухожилия становятся менее утомляемой частью мышцы. Закрепляются сухожилия на костях, проникая волокнами в толщу костной ткани (связь с костью настолько крепка, что скорее разорвется сухожилие, чем оно оторвется от кости). Сухожилия могут переходить на поверхность мышцы и покрывать их на большем или меньшем расстоянии, образуя блестящую оболочку, которая называется сухожильным зеркалом.
В определенных участках в мышцу входят сосуды, ее кровоснабжающие, и нервы, ее иннервирующие. Место вступления их называется воротами органа. Внутри мышцы сосуды и нервы разветвляются по внутреннему перимизию и доходят до его рабочих единиц - мышечных волокон, на которых сосуды образуют сети капилляров, а нервы разветвляются на:
1) чувствительные волокна - идут от чувствительных нервных окончаний проприорецепторов, расположенных во всех участках мышц и сухожилий, и выносят импульс, направляющийся через клетку спинального ганглия в мозг;
2) двигательные нервные волокна, проводящие импульс от мозга:
а) к мышечным волокнам, заканчиваются на каждом мышечном волокне особой моторной бляшкой,
б) к сосудам мышц - симпатические волокна, несущие импульс от мозга через клетку симпатического ганглия к гладким мышцам сосудов,
в) трофические волокна, заканчивающиеся на соединительнотканной основе мышцы. Поскольку рабочей единицей мышц является мышечное волокно, то именно их количество определяет
силу мышцы; не от длины мышечных волокон, а от количества их в мышце зависит сила мышцы. Чем больше мышечных волокон в мышце, тем она сильнее. При сокращении мышца укорачивается на половину своей длины. Чтобы подсчитать количество мышечных волокон, делают разрез перпендикулярно их продольной оси; полученная площадь поперечно перерезанных волокон - это физиологический поперечник. Площадь разреза всей мышцы перпендикулярная ее продольной оси называется анатомическим поперечником. В одной и той же мышце может быть один анатомический и несколько физиологических поперечников, образовавшихся в том случае, если в мышце мышечные волокна короткие и имеют различное направление. Так как сила мышцы зависит от количества мышечных волокон в них, то она выражается отношением анатомического поперечника к физиологическому. В мышечном брюшке имеется всего один анатомический поперечник, а физиологических может быть различное количество (1:2, 1:3, ..., 1:10 и т. д.). Большое количество физиологических поперечников свидетельствует о силе мышцы.
Мышцы бывают светлые и темные. Цвет их зависит от функции, строения и кровенаполнения. Темные мышцы богаты миоглобином (миогематином) и саркоплазмой, они более выносливые. Светлые мышцы беднее этими элементами, они более сильные, но менее выносливые. У разных животных, в различном возрасте и даже в разных участках тела цвет мышц бывает различен: у лошадей мышцы темнее, чем у других видов животных; у молодняка светлее, чем у взрослых; на конечностях темнее, чем на теле.
КЛАССИФИКАЦИЯ МЫШЦ
Каждая мышца является самостоятельным органом и имеет определенную форму, величину, строение, функцию, происхождение и положение в организме. В зависимости от этого все скелетные мышцы подразделяются на группы.
Внутренняя структура мышцы.
Скелетные мышцы по взаимоотношениям мышечных пучков с внутримышечными соединительнотканными образованиями могут иметь самое различное строение, что, в свою очередь, обусловливает их функциональные различия. О силе мышц принято судить по количеству мышечных пучков, определяющих величину физиологического поперечника мышцы. Отношение физиологического поперечника к анатомическому, т.е. соотношение площади поперечного сечения мышечных пучков к наибольшей площади поперечного сечения мышечного брюшка, дает возможность судить о степени выраженности ее динамических и статических свойств. Различия в этих соотношениях позволяют подразделять скелетные мышцы на динамические, динамо– статические, статодинамические и статические.
Проще всего построены простые динамические мышцы . В них нежный перимизий, мышечные волокна длинные, идут вдоль продольной оси мышцы или под некоторым углом к ней, в связи с чем анатомический поперечник совпадает с физиологическим 1:1. Эти мышцы обычно связаны больше с динамической нагрузкой. Обладая большой амплитудой: они обеспечивают большой размах движения, но сила их небольшая – эти мышцы относятся к быстрым, ловким, но и быстро утомляющимся.
Статодинамические мышцы имеют более сильно развитый перимизий (и внутренний и наружный) и более короткие мышечные волокна, идущие в мышцах в различных направлениях, т. е. образующие уже
Классификация мышц: 1 – односуставные, 2 – двусуставные, 3 – многосуставные, 4 – мышцы–связки.
Типы строения статодинамических мышц: а – одноперистая, б – двуперистая, в – многоперистая, 1 – сухожилия мышц, 2 – пучки мышечных волокон, 3 – сухожильные прослойки, 4 – анатомический поперечник, 5 – физиологический поперечник.
множество физиологических поперечников. По отношению к одному общему анатомическому поперечнику в мышце может оказаться 2, 3, 10 физиологических поперечников (1:2, 1:3, 1:10), что дает основание говорить о том, что статодинамические мышцы сильнее динамических.
Статодинамические мышцы выполняют в большей мере статическую функцию во время опоры, удерживая разогнутыми суставы при стоянии животного, когда под действием массы тела суставы конечностей стремятся согнуться. Вся мышца может быть пронизана сухожильным тяжем, который дает возможность во время статической работы выполнять роль связки, снимая нагрузку с мышечных волокон и становясь мышечным фиксатором (двуглавая мышца у лошадей). Для этих мышц характерна большая сила и значительная выносливость.
Статические мышцы могут развиться в результате большой статической нагрузки, падающей на них. Мышцы, подвергшиеся глубокой перестройке и почти полностью утратившие мышечные волокна, фактически превращается в связки, которые способны выполнять лишь статическую функцию. Чем ниже на теле расположены мышцы, тем более они статичны по структуре. Они выполняют большую статическую работу при стоянии и опоре конечности о почву во время движения, закрепляя суставы в определенном положении.
Характеристика мышц по действию.
Согласно функции каждая мышца обязательно имеет два пункта закрепления на костных рычагах - головкой и сухожильным окончанием - хвостом, или апоневрозом. В работе один из этих пунктов будет неподвижной точкой опоры - punctum fixum , второй - подвижной -punctum mobile. У большинства мышц, особенно конечностей, эти пункты меняются в зависимости от выполняемой функции и местонахождения точки опоры. Мышца, закрепленная на двух пунктах (голове и плече), может двигать головой, когда неподвижная точка опоры ее на плече, и, наоборот, будет двигать плечом, если во время движенияpunctum fixum этой мышцы будет на голове.
Мышцы могут действовать только на один или два сустава, но чаще они являются многосуставными. Каждая ось движения на конечностях обязательно имеет две группы мышц с противоположным действием.
При движении по одной оси обязательно будут мышцы-сгибатели -флексоры и разгибатели -экстензоры , в некоторых суставах возможно приведение -аддукция , отведение -абдукция или вращение -ротация , причем вращение в медиальную сторону называетсяпронацией , а вращение наружу в латеральную сторону -супинацией .
Выделяются еще мышцы - напрягатели фасций - тензоры . Но при этом обязательно надо помнить, что в зависимости от характера нагрузки одна и та же
многосуставная мышца может работать как флексор одного сустава или как экстензор другого сустава. Примером может быть двуглавая мышца плеча, которая может оказывать действие на два сустава - плечевой и локтевой (закрепляется на лопатке, перебрасывается через вершину плечевого сустава, проходит внутри угла локтевого сустава и закрепляется на лучевой кости). При висячей конечности punctum fixum у двуглавой мышцы плеча будет в области лопатки, в этом случае мышца тянет вперед, лучевую кость и локтевой сустав сгибает. При опоре конечности о почвуpunctum fixum находится в области конечного сухожилия на лучевой кости; мышца работает уже как экстензор плечевого сустава (удерживает плечевой сустав в разогнутом состоянии).
Если мышцы оказывают противоположное действие на сустав, они называются антагонистами . Если их действие осуществляется в одном направлении, они называются «сотоварищами» -синергистами . Все мышцы, сгибающие один и тот же сустав, будут синергистами, экстензоры этого сустава по отношению к флексорам будут антагонистами.
Вокруг естественных отверстий расположены мышцы–запиратели -сфинктеры , для которых характерно круговое направление мышечных волокон;констрикторы , или суживатели, которые также

относятся к типу круглых мышц, но имеют иную форму; дилататоры , или расширители, при сокращении открывают естественные отверстия.
По анатомическому строению мышцы делятся в зависимости от количества внутримышечных сухожильных прослоек и направления мышечных прослоек:
одноперистые - для них характерно отсутствие сухожильных прослоек и мышечные волокна присоединяются к сухожилию одной стороны;
двуперистые - для них характерно наличие одной сухожильной прослойки и мышечные волокна присоединяются к сухожилию с двух сторон;
многоперистые - для них характерно наличие двух и более сухожильных прослоек, в результате этого мышечные пучки сложно переплетаются и к сухожилию подходят с нескольких сторон.
Классификация мышц по форме
Среди огромного многообразия мышц по форме можно выделить условно следующие основные типы: 1) Длинные мышцы соответствуют длинным рычагам движения и поэтому встречаются главным образом на конечностях. Имеют веретенообразную форму, средняя часть называется брюшком, конец, соответствующий началу мышцы, - головкой, противоположный конец - хвостом. Сухожилие длинных мышц имеет форму ленты. Некоторые длинные мышцы начинаются несколькими головками (многоглавые)
на различных костях, что усиливает их опору.
2) Короткие мышцы находятся на тех участках тела, где размах движений невелик (между отдельными позвонками, между позвонками и ребрами и т.д.).
3) Плоские (широкие) мышцы располагаются преимущественно на туловище и поясах конечностей. Они имеют расширенное сухожилие, называемое апоневрозом. Плоские мышцы обладают не только двигательной функцией, но также опорной и защитной.
4) Встречаются также и другие формы мышц: квадратная ,круговая ,дельтовидная ,зубчатая ,трапециевидная ,веретеновидная и др.
ВСПОМОГАТЕЛЬНЫЕ ОРГАНЫ МЫШЦ
При работе мышц часто создаются условия, снижающие эффективность их работы, особенно на конечностях, когда направление мышечной силы при сокращении происходит параллельно направлению плеча рычага. (Самое выгодное действие мышечной силы тогда, когда она направлена под прямым углом к плечу рычага.) Однако недостаток этого параллелизма в работе мышц устраняется рядом дополнительных приспособлений. Так, например, в местах приложения силы кости имеют бугры, гребни. Под сухожилия подкладываются специальные косточки (или вправляются между сухожилиями). В местах сочленения кости утолщаются, отделяя мышцу от центра движения в суставе. Одновременно с эволюцией мышечной системы тела развиваются как неотъемлемая ее часть вспомогательные приспособления, улучшающие условия работы мышц и помогающие им. К ним относятся фасции, бурсы, синовиальные влагалища, сесамовидные косточки, специальные блоки.
Вспомогательные органы мышц:
А - фасции в области дистальной трети голени лошади (на поперечном разрезе), Б - удерживатели и синовиальные влагалища сухожилий мышц в области заплюсневого сустава лошади с медиальной поверхности, В - фиброзное и синовиальное влагалища на продольном и В" - поперечном срезах;
I - кожа, 2 - подкожная клетчатка, 3 - поверхностная фасция, 4 - глубокая фасция, 5 собственная фасция мышц, 6 - собственная фасция сухожилия (фиброзное влагалище), 7 - соединения поверхностной фасции с кожей, 8 - межфасциалъные соединения, 8 - сосудисто-нервный пучок, 9 - мышцы, 10 - кость, 11 - синовиальные влагалища, 12 - удерживатели разгибателей, 13 - удерживатели сгибателей, 14 - сухожилие;
а - париетальный и b - висцеральный листки синовиального влагалища, с - брыжейка сухожилия, d - места перехода париетального листка синовиального влагалища в его висцеральный листок, е - полость синовиального влагалища
Фасции.
Каждая мышца, группа мышц и вся мускулатура тела одеты специальными плотными фиброзными оболочками, называемыми фасциями - fasciae . Они плотно притягивают мышцы к скелету, фиксируют их положение, способствуя уточнению направления силы действия мышц и их сухожилий, поэтому хирурги называют их футлярами мышц. Фасции отграничивают мышцы друг от друга, создают опору для мышечного брюшка при его сокращении и устраняют трение мышц друг от друга. Фасции еще называют мягким скелетом (считают остатком перепончатого скелета предков - позвоночных). Они помогают и в опорной функции костного скелета - натяжение фасций при опоре снижает нагрузку на мышцы, смягчает ударную нагрузку. В этом случае фасции берут на себя амортизационную функцию. Они богаты рецепторами и сосудами, в связи с чем вместе с мышцами обеспечивают мышечно–суставное чувство. Весьма существенную роль играют в регенерационных процессах. Так, если при удалении пораженного хрящевого мениска в коленном суставе на его место вживить лоскут фасции, не потерявшей связь с основным ее пластом (сосудами и нервами), то при определенной тренировке через некоторое время на ее месте дифференцируется орган с выполнением функции мениска, работа сустава и конечности в целом восстанавливается. Таким образом, изменяя локальные условия биомеханической нагрузки на фасции, можно их использовать как источник ускоренной регенерации структур опорно–двигательного аппарата при аутопластике хрящевой и костной тканей в восстановительной и реконструктивной хирургии.
С возрастом фасциальные футляры утолщаются, делаются более прочными.
Под кожей туловище покрыто поверхностной фасцией и связано с ней рыхлой соединительной тканью. Поверхностная, или подкожная, фасция - fascia superficialis, s. subcutanea - отделяет кожу от поверхностных мышц. На конечностях она может иметь прикрепления на коже и костных выступах, что способствует через посредство сокращений подкожных мышц осуществлению сотрясений кожного покрова, как это имеет место у лошадей, когда они освобождаются от назойливых насекомых или при стряхивании приставшего к коже мусора.
На голове под кожей расположена поверхностная фасция головы – f. superficialis capitis , в которой заключены мышцы головы.
Шейная фасция – f. cervicalis лежит вентрально в области шеи и прикрывает трахею. Различают фасцию шеи и грудобрюшную фасцию. Каждая из них соединяется друг с другом дорсально вдоль надостистой и выйной связок и вентрально - по срединной линии живота - белой линии -linea alba.
Шейная фасция лежит вентрально, прикрывая трахею. Ее поверхностный лист закрепляется на каменистой части височной кости, подъязычной кости и крае крыла атланта. Она переходит в фасции глотки, гортани и околоушную. Затем идет вдоль длиннейшей мышцы головы, дает межмышечные перегородки в этой области и достигает лестничной мышцы, сливаясь с ее перимизием. Глубокая пластина этой фасции отделяет вентральные мышцы шеи от пищевода и трахеи, закрепляется на межпоперечных мышцах, впереди переходит на фасции головы, а каудально достигает первого ребра и грудины, следуя дальше как внутригрудная фасция.
С шейной фасцией связана шейная подкожная мышца - m. cutaneus colli . Она идет вдоль шеи, ближе к
ее вентральной поверхности и переходит на лицевую поверхность к мышцам рта и нижней губы. Грудопоясничная фасция – f. thoracolubalis лежит дорсально на туловище и закрепляется на остистых
отростках грудных и поясничных позвонков и маклоке. Фасция образует поверхностную и глубокую пластину. Поверхностная закрепляется на маклоке и остистых отростках позвонков поясничного и грудного отделов. В области холки она закрепляется на остистых и поперечных отростках и называется поперечно–остистой фасцией. На ней закрепляются мышцы, идущие на шею и к голове. Глубокая пластина расположена только на пояснице, закрепляется на поперечно–реберных отростках и дает начало некоторым брюшным мышцам.
Грудобрюшная фасция – f. thoracoabdominalis лежит латерально по бокам от грудной и брюшной полости и закрепляется вентрально по белой линии живота –linea alba .
С грудобрюшной поверхностной фасцией связана грудобрюшная, или кожная, мышца туловища - m. cutaneus trunci - довольно обширная по площади с продольно идущими волокнами. Расположена она по бокам от грудной и брюшной стенок. Каудально отдает пучки в коленную складку.
Поверхностная фасция грудной конечности – f. superficialis membri thoracici является продолжением грудобрюшной фасции. Она значительно утолщена в области запястья и формирует фиброзные влагалища для сухожилий мышц, которые здесь проходят.
Поверхностная фасция тазовой конечности – f. superficialis membri pelvini является продолжением грудопоясничной и значительно утолщена в области заплюсны.
Под поверхностной фасцией расположена глубокая, или собственно фасция – fascia profunda . Она окружает конкретные группы мышц–синергистов или отдельные мышцы и, прикрепляя их в определенном положении на костной основе, обеспечивает им оптимальные условия для самостоятельных сокращений и предотвращает их боковые смещения. В отдельных участках тела, где требуется более дифференцированное движение, от глубокой фасции отходят межмышечные связи и межмышечные перегородки, образующие обособленные фасциальные футляры для отдельных мышц, которые часто относят к собственным фасциям(fascia propria). Там, где требуется групповое усилие мышц, межмышечные перегородки отсутствуют и глубокая фасция, приобретая особенно мощное развитие, имеет четко выраженные тяжи. За счет местных утолщений глубокой фасции в области суставов образуются поперечные, или кольцевидной формы, перемычки: сухожильные дуги, удерживатели сухожилий мышц.
В области головы поверхностная фасция делится на следующие глубокие: Лобная фасция идет со лба на спинку носа; височная - по височной мышце; околоушно-жевательная покрывает околоушную слюнную железу и жевательную мышцу; щечная идет в области боковой стенки носа и щеки и подчелюстная - с вентральной стороны между телами нижней челюсти. Щечно-глоточная фасция идет с каудальной части щечной мышцы.
Внутригрудная фасция – f. endothoracica выстилает внутреннюю поверхность грудной полости. Поперечно–брюшная фасция – f. transversalis выстилает внутреннюю поверхность брюшной полости.Тазовая фасция – f. pelvis выстилает внутреннюю поверхность тазовой полости.
В области грудной конечности поверхностная фасция делится на следующие глубокие: фасции лопатки, плеча, предплечья, кисти, пальцев.
В области тазовой конечности поверхностная фасция делится на следующие глубокие: ягодичную (покрывает область крупа), фасции бедра, голени, стопы, пальцев
Во время движения фасции играют важную роль в качестве приспособления для присасывания крови и лимфы из нижележащих органов. С мышечных брюшков фасции переходят на сухожилия, окружают их и закрепляются на костях, удерживая сухожилия в определенном положении. Такой фиброзный футляр в виде трубки, через которую проходят сухожилия, называется фиброзным влагалищем сухожилия - vagina fibrosa tendinis . Фасция в определенных местах может утолщаться, образуя лентообразные кольца вокруг сустава, притягивающие группу сухожилий, перебрасывающихся через него. Их еще называют кольцевыми связками. Эти связки особенно хорошо выражены в области запястья и заплюсны. В отдельных местах фасция является местом закрепления мышцы, которая ее напрягает,
В местах большого напряжения, особенно при статической работе, фасции утолщаются, волокна их приобретают различное направление, не только способствуя укреплению конечности, но и выполняя роль пружинящего, амортизационного приспособления.
Бурсы и синовиальные влагалища.
Для того чтобы предотвратить трение мышц, сухожилий или связок, смягчить их соприкосновение с другими органами (костью, кожей и т. д.), облегчить скольжение при больших размахах движения, между листами фасций образуются щели, выстланные оболочкой, выделяющей в образовавшуюся полость слизь или синовию, в зависимости от чего различают синовиальные и слизистые бурсы. Слизистые бурсы – bursa mucosa – (изолированные «мешочки»), образованные в уязвимых местах под связками, называются подсвязочными, под мышцами - подмышечными, под сухожилиями - подсухожильными, под кожей - подкожными. Полость их заполнена слизью и они могут быть постоянными или временными (мозоли).
Бурса, которая образуется за счет стенки капсулы сустава, благодаря чему ее полость сообщается с полостью сустава, называется синовиальной бурсой - bursa synovialis . Такие бурсы заполнены синовией и расположены главным образом в областях локтевого и коленного суставов, и их поражение угрожает суставу – воспаление этих бурс вследствие травмы может привести к артриту, поэтому в дифференциальной диагностике знание расположения и строении синовиальных бурс необходимо, оно определяет лечение и прогноз болезни.
Несколько сложнее построены синовиальные влагалища сухожилий –vagina synovialis tendinis , в которых проходят длинные сухожилия, перебрасываясь через запястный, заплюсневый и путовый суставы. Синовиальное влагалище сухожилий отличается от синовиальной сумки тем, что имеет гораздо большие размеры (длину, ширину) и двойную стенку. Оно полностью охватывает движущееся в нем сухожилие мышцы, вследствие этого синовиальное влагалище не только выполняет функцию бурсы, но и укрепляет положение сухожилия мышцы на значительном ее протяжении.

Подкожные бурсы лошади:
1 - подкожная затылочная бурса, 2 - подкожная париетальная бурса; 3 - подкожная скуловая бурса, 4 - подкожная бурса угла нижней челюсти; 5 - подкожная предгрудинная бурса; 6 - подкожная локтевая бурса; 7 - подкожная латеральная бурса локтевого сустава, 8 - подсвязочная бурса локтевого разгибателя запястья; 9 - подкожная бурса абдуктора первого пальца, 10 - медиальная подкожная бурса запястья; 11 - подкожная предкарпальная бурса; 12 - латеральная подкожная бурса; 13 - пальмарная (статарная) подкожная пальцевая бурса; 14 - подкожная бурса четвертой пястной кости; 15, 15" - медиальная и латеральная подкожные бурсы лодыжки; /6 - подкожная пяточная бурса; 17 - подкожная бурса большеберцовой шероховатости; 18, 18" - подфасциальная подкожная предколенная бурса; 19 - подкожная седалищная бурса; 20 - подкожная вертлужная бурса; 21 - подкожная бурса крестца; 22, 22" - подфасциальная подкожная бурса маклока; 23, 23" - подкожная подсвязочная бурса надостистой связки; 24 - подкожная предлопаточная бурса; 25, 25" - подсвязочные каудальная и краниальная бурсы выйной связки
Синовиальные влагалища образуются внутри фиброзных влагалищ, закрепляющих длинные сухожилия мышц при их прохождении через суставы. Внутри стенка фиброзного влагалища выстилается синовиальной оболочкой, образуя париетальный (наружный) лист этой оболочки. Сухожилие, проходящее через этот участок, тоже покрыто синовиальной оболочкой, еевисцеральным (внутренним) листом . Скольжение во время движения сухожилия происходит между двумя листками синовиальной оболочки и синовии, находящейся между этими листками. Два листка синовиальной оболочки связаны между собой тонкой двухслойной и короткой брыжейкой - переходом париентального листа в висцеральный. Синовиальное влагалище, таким образом, представляет собой тончайшую двухслойную замкнутую трубочку, между стенками которой находится синовиальная жидкость, способствующая скольжению в ней длинного сухожилия. При травмах в области суставов, где имеются синовиальные влагалища, приходится дифференцировать источники выделяющейся синовии, выясняя, вытекает она из сустава или синовиального влагалища.
Блоки и сесамовидные кости.
Способствуют улучшению условии работы мышц блоки и сесамовидные косточки. Блоки – trochlea – это определенной формы участки эпифизов трубчатых костей, через которые перекидываются мышцы. Они представляет собой костный выступ и желобок в нем, где проходит сухожилие мышц, благодаря чему сухожилия не смещаются в сторону и увеличивается рычаг приложения силы. Блоки образуются там, где требуется изменение направления действия мышцы. Они покрыты гиалиновым хрящом, улучшающим скольжение мышцы, здесь же нередко имеются синовиальные сумки или синовиальные влагалища. Блоки имеют плечевая и бедренная кости.
Сезамовидные кости – ossa sesamoidea – представляют собой костные образования, которые могут образовываться как внутри сухожилий мышц, так и в стенке капсулы сустава. Они формируются в области очень сильного напряжения мышц и обнаруживаются в толще сухожилий. Располагаются сесамовидные кости или на вершине сустава, или на выступающих краях сочленяющихся костей, или там, где требуется создать подобие мышечного блока, чтобы изменить направление усилий мышцы при ее сокращении. Они изменяют угол прикрепления мышц и тем самым улучшают условия их работы, уменьшая трение. Иногда их называют «окостеневшими участками сухожилий», но необходимо помнить, что они проходят только две стадии развития (соединительнотканную и костную).
Самая крупная сесамовидная кость - коленная чашечка - patella вправлена в сухожилия четырехглавой мышцы бедра и скользит по надмыщелкам бедренной кости. Более мелкие сесамовидные косточки расположены под сухожилиями пальцевых сгибателей с пальмарной и плантарной сторон путового (по две на каждый) сустава. Со стороны сустава эти косточки покрыты гиалиновым хрящом.