Гладка м'язова тканина: особливості будови. Властивості гладкої м'язової тканини. Гладкі та поперечно-смугасті м'язові тканини
1. Встановіть відповідність між залізою та характеристикою, яка їй відповідає. Для цього до кожного елемента першого стовпця підберіть позицію другого стовпця. Впишіть у таблицю цифри вибраних відповідей.
^ ХАРАКТЕРИСТИКА ЗАЛІЗУ
А) недолік гормону, що виробляється, викликає цукровий діабет 1) наднирник
Б) виробляє гормон інсулін; 2) підшлункова.
В) заліза змішаної секреції
г) виробляє гормон інсулін
Д) складається з кіркового та мозкового шару
Е) заліза парна
| А | Б | У | Г | Д | Е |
2. Встановіть відповідність між ознакою та форменим елементом, до якого ця ознака відноситься. Для цього до кожного елемента першого стовпця підберіть позицію другого стовпця. Впишіть у таблицю цифри вибраних відповідей.
^ ОЗНАК ФОРМОВИЙ ЕЛЕМЕНТ
А) має ядро всіх стадіях розвитку 1) еритроцит
Б) у зрілому стані ядра не має 2) лейкоцит
В) здатний до фагоцитозу
Г) здатний до самостійного руху
Д) містить гемоглобін
Г) надає крові червоного кольору
| А | Б | У | Г | Д | Е |
3. Встановіть відповідність між характеристикою кола кровообігу та його назвою. Для цього до кожного елемента першого стовпця підберіть позицію другого стовпця. Впишіть у таблицю цифри вибраних відповідей.
^ ХАРАКТЕРИСТИКА КРУГА КРОВООБІГУ НАЗВА
А) починається у лівому шлуночку 1) велике коло
Б) кров тече у легені 2) малий круг
В) кров артеріальна перетворюється на венозну
Г) закінчується у лівому передсерді
Д) кров виходить із серця під тиском 30 мм.рт.ст.
Е) кров виходить із серця під тиском 120 мм.рт.ст.
| А | Б | У | Г | Д | Е |
4. Визначте правильну послідовність проходження кисню повітря з атмосфери у клітини. У відповіді напишіть відповідну послідовність літер.
А) трахея Б) кров В) бронхи Г) тканини Д) альвеоли легень
 На малюнку зображено серце людини. Покажи, де розташований правий шлуночок серця; чому стінки шлуночків мають різну товщину?
На малюнку зображено серце людини. Покажи, де розташований правий шлуночок серця; чому стінки шлуночків мають різну товщину?
6.  Покажи на малюнку тім'яну кістку, до якого відділу черепа вона належить? Який відділ черепа у людини розвинений краще та чому?
Покажи на малюнку тім'яну кістку, до якого відділу черепа вона належить? Який відділ черепа у людини розвинений краще та чому?
7.  Покажи на малюнку променеву кістку людини. Як змінилася верхня кінцівка людини у зв'язку з прямоходінням та трудовою діяльністю?
Покажи на малюнку променеву кістку людини. Як змінилася верхня кінцівка людини у зв'язку з прямоходінням та трудовою діяльністю?
8. На малюнку зображена система травлення людини, покажи і назви орган, що виробляє травний сік і гормони одночасно.
9. 
 Як змінився пояс нижніх кінцівок та нижня кінцівка людини у зв'язку з прямоходінням?
Як змінився пояс нижніх кінцівок та нижня кінцівка людини у зв'язку з прямоходінням?
10. 
Що зображено малюнку? Розкажи про особливості цих кісток людини.
11. Розкажи про особливості будови хребта людини у зв'язку з прямоходінням.
Розкажи про особливості будови хребта людини у зв'язку з прямоходінням.
12.  Назви судини, зображені на малюнку, розкажи про особливості їх будови та функції.
Назви судини, зображені на малюнку, розкажи про особливості їх будови та функції.
13.  Розкажи, як надати першу допомогу постраждалим.
Розкажи, як надати першу допомогу постраждалим.
14. Між позиціями першого та другого стовпців наведеної нижче таблиці є певний зв'язок.
Яке поняття слід вписати на місце пропуску у цій таблиці? 1) грудина; 2) слізна залоза; 3) гіпофіз; 4) печінка. Що таке ендокринна система?
15. Вибери три відповіді. До ознак нервової тканини відносять:
А) тканина утворена клітинами, що мають тіло та відростки
Б) клітини здатні скорочуватися
В) між клітинами є контакти, які називають синапсами
Г) клітинам властива збудливість
Д) між клітинами багато міжклітинної речовини
Які властивості мають гладкі і поперечно-смугасті м'язи і яким відділом нервової системи регулюється кожен з них?
Що відбувається при гіподинамії та систематичних фізичних навантаженнях?
До якого типу тканин відноситься кров і чому?
У чому заслуга Луї Пастера та Іллі Ілліча Мечникова?
Що дало людству відкриття імунітету?
Яке значення вакцин та лікувальних сироваток? Чим вони відрізняються?
Чому при переливанні крові слід враховувати групи крові донора та реципієнта? У яких випадках слід враховувати резус-фактор?
Як запобігти хворобам серцево-судинної системи? Що необхідно робити для зміцнення серцево-судинної системи?
Як впливає куріння тютюну на дихальну та кровоносну системи?
Чим клінічна смерть відрізняється від біологічної?
Як змінюється їжа в ротовій порожнині та в шлунку? Які властивості мають ферменти?
Як змінюється їжа в дванадцятипалій кишці?
Які процеси відбуваються у тонкій та товстій кишці? Що відбувається при дисбактеріозі?
Яке значення мають вітаміни? Як зберегти вітаміни в їжі?
Розгляньте схему сечовидільної системи та опишіть будову та функції нирок, сечоводів, сечового міхура та сечівника. Як функціонує нефрон?
Важливим властивістю гладкого м'язає її велика пластичність, тобто здатність зберігати надану розтягуванням довжину без зміни напруги. Відмінність між скелетним м'язом, що володіє малою пластичністю, і гладким м'язом з добре вираженою пластичністю, легко виявляється, якщо їх спочатку повільно розтягнути, а потім зняти вантаж, що розтягує. відразу ж коротшає після зняття вантажу. На відміну від цього гладкий м'яз після зняття вантажу залишається розтягнутим доти, поки під впливом будь-якого подразнення не виникає його активного скорочення.
Властивість пластичності має дуже велике значення для нормальної діяльності гладких м'язів стінок порожнистих органів, наприклад сечового міхура: завдяки пластичності гладкої мускулатури стінок міхура тиск усередині нього відносно мало змінюється при різному ступені наповнення.
Збудливість та збудження
Гладкі м'язименш збудливі, ніж скелетні: їх пороги подразнення вищі, а хронаксія довша. Потенціали дії більшості гладком'язових волокон мають малу амплітуду (порядку 60 мв замість 120 у скелетних м'язових волокнах) і більшу тривалість - до 1-3 секунд. на Рис. 151показаний потенціал дії одиночного волокна м'яза матки.
Рефрактерний період продовжується протягом усього періоду потенціалу дії, тобто 1-3 секунди. Швидкість проведення збудження варіює у різних волокнах від кількох міліметрів до кількох сантиметрів на секунду.
Існує велика кількість різних типів гладких м'язів у тілі тварин та людини. Більшість порожнистих органів тіла вистелено гладкими м'язами, що мають сенцитіальний тип будови. Окремі волокна таких м'язів дуже тісно примикають один до одного і складається враження, що вони складають морфологічно єдине ціле.
Протеелектронномікроскопічні дослідження показали, що мембранної та протоплазматичної безперервності між окремими волокнами м'язового синцитію не існує: вони відокремлені один від одного тонкими (200-500 Å) щілинами. Поняття «синцитіальна будова» є нині скоріше фізіологічним, ніж морфологічним.
Синцитій- це функціональне утворення, яке забезпечує те, що потенціали дії та повільні хвилі деполяризації можуть безперешкодно поширюватися з одного волокна на інше. Нервові закінчення розташовані лише на невеликій кількості волокон синцитію. Однак внаслідок безперешкодного поширення збудження з одного волокна на інше залучення в реакцію всього м'яза може відбуватися, якщо нервовий імпульс надходить до невеликої кількості м'язових волокон.
Скорочення гладкого м'яза
При великій силі одиночного подразнення може бути скорочення гладкої м'язи. Прихований період одиночного скорочення цього м'яза значно більше, ніж скелетного м'яза, досягаючи, наприклад, у кишковій мускулатурі кролика 0,25-1 секунди. Тривалість самого скорочення теж велика. Рис. 152): у шлунку кролика вона досягає 5 секунд, а у шлунку жаби - 1 хвилини і більше. Особливо повільно відбувається розслаблення після скорочення. Хвиля скорочення поширюється по гладкій мускулатурі теж дуже повільно, вона проходить лише близько 3 см на секунду. Але ця повільність скорочувальної діяльності гладких м'язів поєднується з великою їх силою. Так, мускулатура шлунка птахів здатна піднімати 1 кг на 1см2 свого поперечного перерізу.
Тонус гладкого м'яза
Внаслідок повільності скорочення гладкий м'яз навіть при рідкісних ритмічних подразненнях (для шлунка жаби достатньо 10-12 подразнень на хвилину) легко переходить у тривалий стан стійкого скорочення, що нагадує тетанус скелетних м'язів. Однак енергетичні витрати при такому стійкому скороченні гладкого м'яза дуже малі, що відрізняє це скорочення від тетануса поперечно-м'язів.
Причини, внаслідок яких гладкі м'язи скорочуються і розслабляються набагато повільніше, ніж скелетні, ще не з'ясовані. Відомо, що міофібрили гладкого м'яза так само, як і скелетного м'яза, складаються з міозину та актину. Однак у гладких м'язах немає поперечної смугастість, немає мембрани Z і вони набагато багатші саркоплазмою. Очевидно, ці особливості структури гладких м'язових хвиль і зумовлюють повільний темп скорочувального процесу. Цьому відповідає відносно низький рівень обміну речовин гладких м'язів.
Автоматія гладких м'язів
Характерною особливістю гладких м'язів, що відрізняє їх від кістякових, є здатність до спонтанної автоматичної діяльності. Спонтанні скорочення можна спостерігати при дослідженні гладких м'язів шлунка, кишок, жовчного міхура, сечоводів та інших гладком'язових органів.
Автоматія гладких м'язів має міогенне походження. Вона властива самим м'язовим волокнам і регулюється нервовими елементами, які перебувають у стінках гладких органів. Міогенна природа автоматії доведена дослідами на смужках м'язів кишкової стінки, звільнених шляхом ретельного препарування від нервових сплетень, що прилягають до неї. Такі смужки, поміщені в тепле розчин Рінгера-Локка, який насичується киснем, здатні здійснювати автоматичні скорочення. При подальшій гістологічній перевірці було виявлено відсутність у цих м'язових смужках нервових клітин.
У гладких м'язових волокнах розрізняють такі спонтанні коливання мембранного потенціалу: 1) повільні хвилі деполяризації з тривалістю циклу близько декількох хвилин і амплітудою близько 20 мв; 2) малі швидкі коливання потенціалу, що передують виникненню потенціалів дії; 3) потенціали дії.
На всі зовнішні впливи гладкий м'яз реагує зміні частоти спонтанної ритміки, наслідком якої є скорочення та розслаблення м'яза. Ефект подразнення гладкої мускулатури кишки залежить від співвідношення між частотою стимуляції і власною частотою спонтанної ритміки: при низькому тонусі - при рідкісних спонтанних потенціалах дії - прикладене роздратування посилює тонус при високому тонусі у відповідь на роздратування виникає розслаблення, так як надмірне що кожен наступний імпульс потрапляє в рефрактерну фазу від попереднього.
До збудливих тканин в організмі людини належать нервова, секреторна та м'язова тканини. Однак остання відрізняється від інших унікальною властивістю скоротливості завдяки наявності у клітинній структурі мікрофіламентів із спеціалізованих білків – міозину, актину, тропоміозину.
За рахунок цього забезпечуються підтримка пози людини, переміщення в просторі, просування харчової грудки шлунково-кишковим трактом, кровообіг і багато іншого. Залежно від гістологічних особливостей, виконуваних функцій та походження існує класифікація на гладкі та поперечно-смугасті м'язові тканини, також за особливості деякі автори виділяють у третій серцевий підвид. Однак слід розуміти, скорочувальні елементи становлять лише основу цих тканин, і вони не могли б повноцінно працювати без густої мережі кровоносних судин, що забезпечують доставку великої кількості кисню, захисної та енергетично значущої оболонки – сарколеми, а також підкріплюючої пухкої неоформленої
Поперечно-смугаста
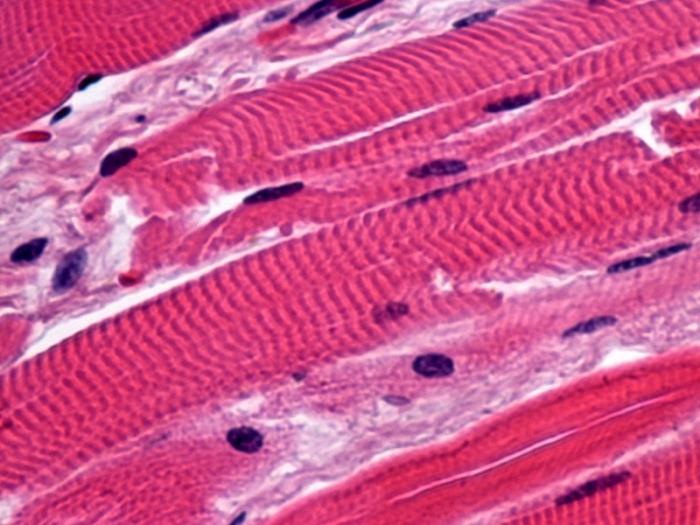
Поперечно-смугасті м'язові тканини, в основному, складають всю скелетну мускулатуру і забезпечують рух суглобів та підтримання пози. Структурно-функціональною одиницею їх є саркомер, а складаються вони, своєю чергою, з миосимпластов - волокон, які утворилися при злитті кількох окремих клітин у процесі диференціювання. На гістологічному препараті поперечно-смугасті м'язові тканини легко відрізнити по багатоядерності та смугастість, чому вони і отримали свою назву. Інший важливою функцією їх є теплоутворення, у зв'язку з чим при зниженні температури у людини виникає тремтіння. Також поперечно-смугаста утворює і структуру міокарда, тільки кардіоміоцити відрізняються відсутністю симпластів. При мікроскопуванні вони є одноядерними веретеноподібними клітинами. Поділяються вони відповідно до своїх функцій на робітники (переважають за кількістю), провідні та секреторні. Саме завдяки другим, серцеві поперечно-смугасті м'язові тканини мають властивість автоматизму, тобто мають здатність самостійно скорочуватися, що й забезпечує безперервну роботу серця. Третій тип клітин є місцем синтезу гормоноподібних речовин, зокрема це передсердний натрійуретичний чинник, сприяє посиленню діурезу.
Гладка
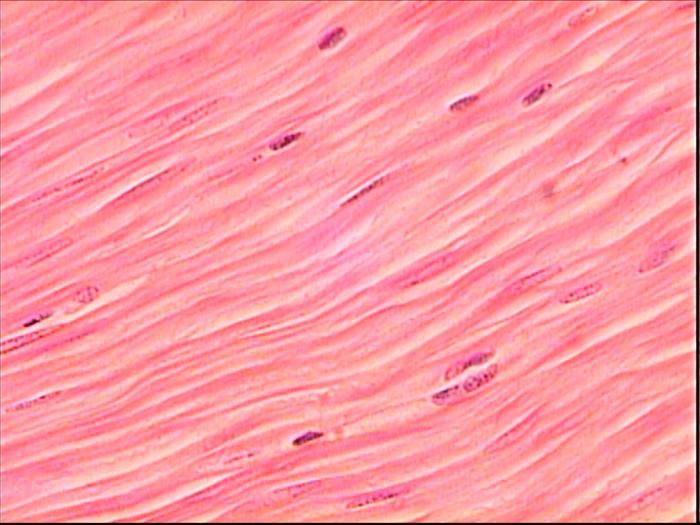
Якщо забезпечує рух м'язова тканина людини - поперечно-смугаста, то перистальтику шлунково-кишкового і сечостатевого тракту, скорочення судинної та бронхіальної стінки забезпечує гладка мускулатура. Вона відрізняється ритмічністю, відносною повільністю, високою мірою розтяжністю та регенераторними здібностями, а також вегетативною іннервацією. Це витягнутої форми одноядерні клітини, без смугастість, і з великою кількістю актину і колагену в структурі. Кожен такий міоцит покритий тонкою базальною мембраною, а групи – ендомізією з пухкої неоформленої сполучної тканини.
Гладкі м'язи представлені в порожнистих органах, кровоносних судинах та шкірі. Гладкі м'язові волокна немає поперечної смугастість. Клітини коротшають внаслідок відносного ковзання ниток. Швидкість ковзання та швидкість розщеплення аденозинтрифосфату в 100-1000 разів менша, ніж у . Завдяки цьому гладкі м'язи добре пристосовані для тривалого стійкого скорочення без утоми, з меншою витратою енергії.
Гладкі м'язиє складовою стінок низки порожнистих внутрішніх органів і беруть участь у забезпеченні функцій, виконуваних цими органами. Зокрема, вони регулюють кровотік у різних органах та тканинах, прохідність бронхів для повітря, переміщення рідин та хімусу (у шлунку, кишечнику, сечоводах, сечовому та жовчному міхурі), скорочення матки при пологах, розмір зіниці, шкірного рельєфу.
Гладком'язові клітини мають веретеноподібну форму, довжину 50-400 мкм, товщину 2-10 мкм (рис. 5.6).
Гладкі м'язи ставляться до мимовільних м'язів, тобто. їх скорочення залежить від волі макроорганізму. Особливості рухової діяльності шлунка, кишечника, кровоносних судин та шкіри певною мірою визначають фізіологічні особливості гладких м'язів цих органів.
Характеристика гладкої мускулатури
- Має автоматизм (вплив інтрамуральної нервової системи носить коригуючий характер)
- Пластичність - здатність довго зберігати довжину без зміни тонусу
- Функціональний синтицій - окремі волокна розділені, але є спеціальні ділянки контакту - нексуси
- Величина потенціалу спокою - 30-50 мВ, амплітуда потенціалу дії менша, ніж у клітин скелетних м'язів
- Мінімальна «критична зона» (збудження виникає, якщо збуджується деяка мінімальна кількість м'язових елементів)
- Для взаємодії актину та міозину необхідний іон Ca 2+ який надходить ззовні
- Тривалість одиночного скорочення велика
Особливість гладких м'язівїх здатність виявляти повільні ритмічні та тривалі тонічні скорочення. Повільні ритмічні скорочення гладких м'язів шлунка, кишечника, сечоводів та інших порожнистих органів сприяють переміщенню їхнього вмісту. Тривалі тонічні скорочення гладких м'язів сфінктерів порожнистих органів перешкоджають довільному виходу їхнього вмісту. Гладкі м'язи стінок кровоносних судин також знаходяться в стані постійного тонічного скорочення і впливають на рівень артеріального тиску крові і кровопостачання організму.
Важливою властивістю гладких м'язів є їх містичність,тобто. здатність зберігати викликану розтягуванням чи деформацією форму. Висока пластичність гладких м'язів має значення для нормального функціонування органів. Наприклад, пластичність сечового міхура дозволяє при наповненні сечею профілактувати підвищення в ньому тиску без порушення процесу сечоутворення.
Надмірне розтягування гладких м'язів викликає їхнє скорочення. Це у результаті деполяризації мембран клітин, викликаної їх розтягуванням, тобто. гладкі м'язи мають автоматизмом.
Скорочення, викликане розтягуванням, відіграє у авторегуляції тонусу кровоносних судин, переміщенні вмісту шлунково-кишкового тракту та інших процесах.
Рис. 1. А. Волокно скелетного м'яза, клітина серцевого м'яза, гладка м'язова клітина. Б. Саркомір скелетного м'яза. В. Будова гладкого м'яза. Г. Механограма скелетного м'яза та м'язи серця.
Автоматизм в гладких м'язах обумовлений наявністю в них спеціальних пейсмекерних (що задають ритм) клітин. За своєю структурою вони ідентичні іншим гладком'язовим клітинам, але мають особливі електрофізіологічні властивості. У цих клітинах виникають пейсмекерні потенціали, які деполяризують мембрану до критичного рівня.
Порушення гладких клітин викликає збільшення входу іонів кальцію в клітину і вивільнення цих іонів з саркоплазматичного ретикулуму. Внаслідок підвищення концентрації іонів кальцію в саркоплазмі активуються скорочувальні структури, але механізм активації їх у гладкому волокні відрізняється від механізму активації у поперечно-смугастих м'язах. У гладенькій клітці кальцій взаємодієте білком кальмодуліном, який активує легкі ланцюги міозину. Вони з'єднуються з активними центрами актину в протофібрил і роблять «гребінець». Гладкі м'язи розслаблюються пасивно.
Гладкі м'язи відносяться до мимовільних, і їх залежить від волі тварини.
Фізіологічні властивості та особливості гладких м'язів
Гладкі м'язи, так само, як і скелетні, мають збудливість, провідність і скоротливість. На відміну від скелетних м'язів, що мають еластичність, гладкі м'язи мають пластичність - здатність тривалий час зберігати придану їм при розтягуванні довжину без збільшення напруги. Така властивість важлива для виконання функції депонування їжі в шлунку або рідин у жовчному та сечовому міхурі.
Особливості збудливості гладких клітин в певній мірі пов'язані з низькою різницею потенціалів на мембрані в спокої (E 0 = (-30) - (-70) мВ). Гладкі міоцити можуть мати автоматію і мимовільно генерувати потенціал дії. Такі клітини - водії ритму скорочення гладких м'язів є у стінках кишечника, венозних та лімфатичних судин.

Рис. 2. Будова гладком'язової клітини (A. Guyton, J. Hall, 2006)
Тривалість ПД гладких міоцитів може досягати десятків мілісекунд, оскільки ПД у них розвивається переважно за рахунок входу іонів Са 2+ у саркоплазму з міжклітинної рідини через повільні кальцієві канали.
Швидкість проведення ПД мембраною гладких міоцитів мала - 2-10 см/с. На відміну від скелетних м'язів збудження може передаватися з одного гладкого міоциту на інші, що лежать поруч. Така передача відбувається завдяки наявності між гладком'язовими клітинами нексусів, які мають малий опір електричного струму і забезпечують обмін між клітинами іонів Са 2+ та іншими молекулами. В результаті цього гладкий м'яз виявляє властивості функціонального синтицію.
Скоротимість гладких клітин відрізняється тривалим латентним періодом (0,25-1,00 с) і великою тривалістю (до 1 хв) одиночного скорочення. Гладкі м'язи розвивають малу силу скорочення, але здатні довго перебувати у тонічному скороченні без розвитку втоми. Це пов'язано з тим, що на підтримання тонічного скорочення гладкий м'яз витрачає в 100-500 разів менше енергії, ніж скелетний. Тому запаси АТФ, що витрачаються гладким м'язом, встигають відновлюватися навіть під час скорочення і гладкі м'язи деяких структур організму практично постійно перебувають у стані тонічного скорочення. Абсолютна сила гладкого м'яза становить близько 1 кг/см 2 .
Механізм скорочення гладкого м'яза
Найважливішою особливістю гладких клітин є те, що вони збуджуються під впливом численних подразників. у природних умовах ініціюється лише нервовим імпульсом, що приходить до . Скорочення гладкого м'яза може бути викликане як впливом нервових імпульсів, так і дією гормонів, нейромедіаторів, простагландинів, деяких метаболітів, а також впливом фізичних факторів, наприклад розтягуванням. Крім того, збудження та скорочення гладких міоцитів може статися спонтанно – за рахунок автоматик.
Здатність гладких м'язів відповідати скороченням на дію різноманітних факторів створить значні труднощі для корекції порушень тонусу цих м'язів у медичній практиці. Це видно на прикладах труднощів лікування бронхіальної астми, артеріальної гіпертензії, спастичного коліту та інших захворювань, що потребують корекції скорочувальної активності гладких м'язів.
У молекулярному механізмі скорочення гладкого м'яза також є низка відмінностей від механізму скорочення скелетного м'яза. Нитки актину і міозину в гладком'язових клітинах розташовуються менше впорядкування, ніж у скелетних, і тому гладкий м'яз не має поперечної смугастість. В актинових нитках гладкого м'яза немає білка тропоніну і центри актину завжди відкриті для взаємодії з головками міозину. У той же час, головки міозину в стані спокою не енергізовані. Для того щоб відбулася взаємодія актину та міозину, необхідно фосфорилювати головки міозину та надати їм надлишок енергії. Взаємодія актину та міозину супроводжується поворотом головок міозину, при якому актинові нитки втягуються між міозиновими та відбувається скорочення гладкого міоциту.
Фосфорилювання головок міозину проводиться за участю ферменту кінази легких ланцюгів міозину, а дефосфорилювання – за допомогою фосфатази. Якщо активність фосфатази міозину переважає над активністю кінази, то головки міозину дефосфорилуються, зв'язок міозину та актину розривається і розслабляється м'яз.
Отже, щоб відбулося скорочення гладкого міоциту, необхідно підвищити активність кінази легких ланцюгів міозину. Її активність регулюється рівнем іонів Са 2+ у саркоплазмі. Нейромедіатори (ацетилхолін, норадрсналін) або гормони (вазопресин, окситоцин, адреналін) стимулюють свій специфічний рецептор, викликаючи дисоціацію G-білка, а-субодиниця якого далі активує фермент фосфоліпазу С. Фосфолігтза С каталізує утворення іноз мембрани клітини. ІФЗ дифундує до ендоплазматичного ретикулуму і після взаємодії зі своїми рецепторами викликає відкриття кальцієвих каналів та вивільнення іонів Са 2+ з депо до цитоплазми. Збільшення вмісту іонів Са 2+ у цитоплазмі є ключовою подією для ініціації скорочення гладкого міоциту. Збільшення вмісту іонів Са 2+ у саркоплазмі досягається також за рахунок його надходження до міоциту з позаклітинного середовища (рис. 3).
Іони Са 2+ утворюють комплекс з білком кальмодуліном, і комплекс Са 2+ -кальмодулін підвищує кіназну активність легких ланцюгів міозину.
Послідовність процесів, що призводять до розвитку скорочення гладкого м'яза, можна описати наступним чином: вхід іонів Са 2+ в саркоплазму - активація кальмодуліну (шляхом утворення комплексу 4Са 2 -кальмодулін) - активація кінази легких ланцюгів міозину - фосфорилювання головок міозину - зв'язування головок і поворот головок, у якому нитки актина втягуються між нитками міозину — скорочення.

Рис. 3. Шляхи надходження іонів Са 2+ у саркоплазму гладком'язової клітини (а) та видалення їх із саркоплазми (б)
Умови, необхідні для розслаблення гладкого м'яза:
- зниження (до 10-7 М/л і менше) вмісту іонів Са2+ у саркоплазмі;
- розпад комплексу 4Са 2+ -кальмодулін, що призводить до зниження активності кінази легких ланцюгів міозину - дефосфорилювання головок міозину під впливом фосфатази, що призводить до розриву зв'язків ниток актину та міозину.
У умовах еластичні сили викликають щодо повільне відновлення вихідної довжини гладком'язового волокна та її розслаблення.
лекція № 4. Фізіологія м'язів
1. Фізичні та фізіологічні властивості скелетних, серцевих та гладких м'язів
За морфологічними ознаками виділяють три групи м'язів:
1) поперечно-смугасті м'язи (скелетні м'язи);
2) гладкі м'язи;
3) серцевий м'яз (або міокард).
Функції поперечно-смугастих м'язів:
1) рухова (динамічна та статична);
2) забезпечення дихання;
3) мімічна;
4) рецепторна;
5) депонуючий;
6) терморегуляторна.
Функції гладких м'язів:
1) підтримання тиску в порожнистих органах;
2) регуляція тиску у кровоносних судинах;
3) спорожнення порожнистих органів та просування їх вмісту.
Функція серцевого м'яза- Насосна, забезпечення руху крові по судинах.
1) збудливість (нижче, ніж у нервовому волокні, що пояснюється низькою величиною мембранного потенціалу);
2) низька провідність, близько 10-13 м/с;
3) рефрактерність (займає за часом більший відрізок, ніж у нервового волокна);
4) лабільність;
5) скоротливість (здатність укорочуватися чи розвивати напругу).
Розрізняють два види скорочення:
а) ізотоничне скорочення (змінюється довжина, тонус не змінюється);
б) ізометричне скорочення (змінюється тонус без зміни довжини волокна). Розрізняють поодинокі та титанічні скорочення. Поодинокі скорочення виникають при дії одиночного подразнення, а титанічні виникають у відповідь серію нервових імпульсів;
6) еластичність (здатність розвивати напругу при розтягуванні).
Гладкі м'язи мають ті ж фізіологічні властивості, що і скелетні м'язи, але мають свої особливості:
1) нестабільний мембранний потенціал, який підтримує м'язи у стані постійного часткового скорочення – тонусу;
2) мимовільну автоматичну активність;
3) скорочення у відповідь розтяг;
4) пластичність (зменшення розтягування зі збільшенням розтягування);
5) високу чутливість до хімічних речовин.
Фізіологічною особливістю серцевого м'яза є її автоматизм . Порушення виникає періодично під впливом процесів, що протікають у самому м'язі. Здатність до автоматизму мають певні атипові м'язові ділянки міокарда, бідні на міофібрили і багаті на саркоплазму.
2. Механізми м'язового скорочення
Електрохімічний етап м'язового скорочення.
1. Генерація потенціалу дії. Передача збудження на м'язове волокно відбувається за допомогою ацетилхоліну. Взаємодія ацетилхоліну (АХ) з холінорецепторами призводить до їх активації та появи потенціалу дії, що є першим етапом м'язового скорочення.
2. Поширення потенціалу дії. Потенціал дії поширюється всередину м'язового волокна по поперечній системі трубочок, яка є ланкою, що зв'язує між поверхневою мембраною і скоротливим апаратом м'язового волокна.
3. Електрична стимуляція місця контакту призводить до активації ферменту та утворення інозилтрифосфату, який активує кальцієві канали мембран, що призводить до виходу іонів Ca та підвищення їх внутрішньоклітинної концентрації.
Хемомеханічний етап м'язового скорочення.
Теорія хемомеханічного етапу м'язового скорочення була розроблена О. Хакслі у 1954 р. та доповнена у 1963 р. М. Девісом. Основні положення цієї теорії:
1) іони Ca запускають механізм м'язового скорочення;
2) за рахунок іонів Ca відбувається ковзання тонких актинових ниток по відношенню до міозинових.
У спокої, коли іонів Ca мало, ковзання не відбувається, тому що цьому перешкоджають молекули тропоніну та негативно заряди АТФ, АТФ-ази та АДФ. Підвищена концентрація іонів Ca відбувається за рахунок надходження його з міжфібрилярного простору. При цьому відбувається низка реакцій за участю іонів Ca:
1) Ca2+ реагує з трипоніном;
2) Ca2+ активує АТФ-азу;
3) Ca2+ знімає заряди з АДФ, АТФ, АТФ-ази.
Взаємодія іонів Ca з тропоніном призводить до зміни розташування останнього на актиновій нитці, відкриваються активні центри тонкої протофібрили. За рахунок них формуються поперечні містки між актином та міозином, які переміщують актинову нитку у проміжки між міозиновою ниткою. При переміщенні актинової нитки щодо міозинової відбувається скорочення м'язової тканини.
Отже, головну роль у механізмі м'язового скорочення відіграють тропонін білок, який закриває активні центри тонкої протофібрили та іони Ca.
Фізіологія скелетних та гладких м'язів
Лекція 5
У хребетних та людини три види м'язів: поперечносмугасті м'язи скелета, поперечносмугастий м'яз серця - міокард і гладкі м'язи, що утворюють стінки порожнистих внутрішніх органів і судин.
Анатомічною та функціональною одиницею скелетних м'язів є нейромоторна одиниця - руховий нейрон та іннервована ним група м'язових волокон. Імпульси, що посилаються мотонейроном, приводять у дію всі м'язові волокна, що її утворюють.
Скелетні м'язискладаються з великої кількості м'язових волокон. Волокно поперечнополосатого м'яза має витягнуту форму, діаметр його від 10 до 100 мкм, довжина волокна від кількох сантиметрів до 10-12 см. М'язова клітина оточена тонкою мембраною – сарколемою, містить саркоплазму(протоплазму) та численні ядра. Скоротливою частиною м'язового волокна є довгі м'язові нитки. міофібрили, Що складаються в основному з актину, що проходять усередині волокна від одного кінця до іншого, мають поперечну смугастість. Міозин у гладких м'язових клітинах знаходиться в дисперсному стані, але містить багато білка, що відіграє важливу роль у підтримці тривалого тонічного скорочення.
У період відносного спокою скелетні м'язи повністю не розслабляються і зберігають помірний рівень напруги, тобто. м'язовий тонус.
Основні функції м'язової тканини:
1) рухова - забезпечення руху
2) статична – забезпечення фіксації, зокрема й у певної позі
3) рецепторна – у м'язах є рецептори, що дозволяють сприймати власні рухи
4) депонуюча – у м'язах запасаються вода та деякі поживні речовини.
Фізіологічні властивості кістякових м'язів:
Збудливість . Нижче, ніж збудливість нервової тканини. Порушення поширюється вздовж м'язового волокна.
Провідність . Менше провідності нервової тканини.
Рефрактерний період м'язової тканини триваліший, ніж нервової тканини.
Лабільність м'язової тканини значно нижчі, ніж нервової.
Скоротність - Здатність м'язового волокна змінювати свою довжину і ступінь напруги у відповідь на подразнення порогової сили.
При ізотонічному скороченнязмінюється довжина м'язового волокна без зміни тонусу. При ізометричному скороченнязростає напруга м'язового волокна без зміни довжини.
Залежно від умов стимуляції та функціонального стану м'яза може виникнути поодиноке, злите (тетанічне) скорочення або контрактура м'яза.
Поодиноке м'язове скорочення.При подразненні м'яза поодиноким імпульсом струму виникає поодиноке м'язове скорочення.
Амплітуда одиночного скорочення м'яза залежить від кількості міофібрил, що скоротилися в цей момент. Збудливість окремих груп волокон різна, тому порогова сила струму викликає скорочення найбільш збудливих м'язових волокон. Амплітуда такого скорочення мінімальна. При збільшенні сили дратівливого струму процес збудження залучаються і менш збудливі групи м'язових волокон; амплітуда скорочень підсумовується і зростає доти, доки в м'язі не залишиться волокон, не охоплених процесом збудження. У цьому випадку реєструється максимальна амплітуда скорочення, яка не збільшується, незважаючи на подальше наростання сили подразнюючого струму.
Тетанічне скорочення. У природних умовах до м'язових волокон надходять не поодинокі, а ряд нервових імпульсів, на які м'яз відповідає тривалим, тетанічним скороченням, або тетанусом . До тетанічного скорочення здатні лише кістякові м'язи. Гладкі м'язи і поперечносмугастий м'яз серця не здатні до тетанічного скорочення через тривалий рефрактерний період.
Тетанус виникає внаслідок сумування одиночних м'язових скорочень. Щоб виник тетанус, потрібна дія повторних подразнень (або нервових імпульсів) на м'яз ще до того, як закінчиться її одиночне скорочення.
Якщо дратівливі імпульси зближені і кожен із них припадає на той момент, коли м'яз тільки почав розслаблятися, але не встиг ще повністю розслабитися, то виникає зубчастий тип скорочення. зубчастий тетанус ).
Якщо дратівливі імпульси зближені настільки, що кожен наступний припадає на час, коли м'яз ще не встиг перейти до розслаблення від попереднього роздратування, тобто відбувається на висоті її скорочення, виникає тривале безперервне скорочення, що отримала назву гладкого тетанусу .
Гладкий тетанус – нормальний робочий стан скелетних м'язів обумовлюється надходженням із ЦНС нервових імпульсів із частотою 40-50 в 1с.
Зубчастий тетанус виникає при частоті нервових імпульсів до 30 1с. Якщо м'яз отримує 10-20 нервових імпульсів в 1с, він перебуває у стані м'язового тонусу , тобто. помірного ступеня напруги.
Втома м'язів . При тривалому ритмічному подразненні у м'язі розвивається втома. Ознаками його є зниження амплітуди скорочень, збільшення їх латентних періодів, подовження фази розслаблення і, нарешті, відсутність скорочень при подразненні, що триває.
Ще один різновид тривалого скорочення м'язів контрактура. Вона продовжується і при знятті подразника. Контрактура м'язів настає при порушенні обміну речовин або зміні властивостей скоротливих білків м'язової тканини. Причинами контрактури можуть бути отруєння деякими отрутами та лікарськими засобами, порушення обміну речовин, підвищення температури тіла та інші фактори, що призводять до незворотних змін м'язової тканини.
Фізіологічні особливості гладких м'язів.
Гладкі м'язи утворюють стінки (м'язовий шар) внутрішніх органів та кровоносних судин. У міофібрила гладких м'язів немає поперечної смугастість. Це зумовлено хаотичним розташуванням скорочувальних білків. Волокна гладких м'язів відносно коротші.
Гладкі м'язи менш збудливі , Чим поперечносмугасті. Порушення по них поширюється з невеликою швидкістю - 2-15 см/с. Порушення у гладких м'язах може передаватися з одного волокна на інше, на відміну від нервових волокон та волокон поперечносмугастих м'язів.
Скорочення гладкої мускулатури відбувається повільніше і триваліше.
Рефрактерний період у гладких м'язах більш тривалий, ніж у скелетних.
Важливою властивістю гладкого м'яза є його великий пластичність, тобто. здатність зберігати надану розтягуванням довжину без зміни напруги. Ця властивість має істотне значення, оскільки деякі органи черевної порожнини (матка, сечовий міхур, жовчний міхур) іноді значно розтягуються.
Характерною особливістю гладких м'язів є їх здатність до автоматичної діяльностіяка забезпечується нервовими елементами, закладеними в стінках гладком'язових органів.
Адекватним подразником для гладких м'язів є їхнє швидке і сильне розтягування, що має велике значення для функціонування багатьох гладком'язових органів (сечовід, кишечник та інші порожнисті органи)
Особливістю гладких м'язів є також їх висока чутливість до деяких біологічно активних речовин(Ацетилхолін, адреналін, норадреналін, серотонін та ін).
Гладкі м'язи іннервуються симпатичними та парасимпатичними вегетативними нервами, які, як правило, надають протилежний вплив на їхній функціональний стан.
Основні властивості серцевого м'яза.
Стінка серця складається із 3 шарів. Середній шар (міокард) складається з поперечнополосатого м'яза. Серцевий м'яз, як і скелетні м'язи, має властивість збудливості, здатність проводити збудження і скоротливість. До фізіологічних особливостей серцевого м'яза належать подовжений рефрактерний період та автоматизм.
Збудливість серцевого м'яза . Серцевий м'яз менш збудливий, ніж скелетний. Для виникнення збудження в серцевому м'язі необхідний сильніший подразник, ніж для скелетного.
Провідність . Порушення по волокнах серцевого м'яза проводиться з меншою швидкістю, ніж по волокнах скелетного м'яза.
Скоротність . Реакція серцевого м'яза не залежить від сили подразнень, що наносяться. Серцевий м'яз максимально скорочується і на граничне і більш сильне за величиною подразнення.
Рефрактерний період . Серце, на відміну інших збудливих тканин, має значно виражений і подовжений рефрактерний період. Він характеризується різким зниженням збудливості тканини у період її активності. Завдяки цьому серцевий м'яз не здатний до тетанічного (тривалого) скорочення і здійснює свою роботу за типом одиночного м'язового скорочення.
Автоматизм серця . Поза організмом за певних умов серце здатне скорочуватися та розслаблятися, зберігаючи правильний ритм. Здатність серця ритмічно скорочуватися під впливом імпульсів, що у ньому самому, зветься автоматизму.
Класифікація та функції м'язової тканини
Існують 3 види м'язової тканини:
1)поперечно-смугаста скелетна;
2)поперечно-смугаста серцева;
3) гладка.
Функції м'язової тканини.
Поперечно-смугаста скелетна тканина- становить приблизно 40% від загальної маси тіла.
Її функції:
1) динамічна;
2) статична;
3) рецепторна (наприклад, пропріорецептори в сухожиллях - інтрафузальні м'язові волокна (веретеноподібні));
4)депонуюча - вода, мінеральні речовини, кисень, глікоген, фосфати;
5) терморегуляція;
6) емоційні реакції.
Поперечно-смугаста серцева м'язова тканина.
Основна функція- Нагнітальна.
Гладка мускулатура- утворює стінку порожнистих органів та судин.
Її функції:
1) підтримує тиск у порожнистих органах;
2) підтримує величину кров'яного тиску;
3)забезпечує просування вмісту шлунково-кишкового тракту, сечоводів.
Фізіологічні властивості м'язів
Збудливість м'язової тканини (-90 мВ) менше збудливості нервової тканини (-150 мВ).
Провідність м'язової тканини менше провідності нервової тканини, у скелетній тканині (5-6 м/с), а в нервовій – 13 м/с.
Рефрактерність м'язової тканини більше рефрактерності нервової тканини. Для скелетної тканини вона дорівнює 30-40 мс (абсолютна приблизно дорівнює 5 мс, відносна – 30 мс). Рефрактерність гладком'язової тканини дорівнює кількох секунд.
Лабільність м'язової тканини (200-250), нижче за лабільність нервової тканини.
Скоротність виділяють ізотонічне (зміна довжини) та ізометричне (зміна напруги м'язів) скорочення. Ізотонічне скорочення може бути: концентричним (м'яз вкорочується), ексцентричним (довжина м'яза збільшується).
Провідна система м'язового волокна
При нанесенні подразнення на постсинаптичній мембрані м'яза виникає постсинаптичний потенціал, який і генерує потенціал м'язової дії.
Провідний апарат м'яза включає:
1) поверхнева плазматична мембрана;
2) Т-система;
3) саркоплазматичний ретикулум.
Поверхнева плазматична мембрана - Внутрішній шар мембрани, що покриває м'язове волокно. Вона має електрогенні властивості на всьому протязі. Порушення проходить як за безмієліновим волокном.
Т-система - це система поперечних трубочок, є випинання поверхневої плазматичної мембрани вглиб м'язових волокон. Вони проходять між міофібрилами на рівні Z-мембрани.
Саркоплазматичний ретикулум - замкнуті цистерни із Са2+ (у зв'язаному, іонізованому вигляді – 50 %, у вигляді органічних сполук – 50 %).
Тріада - одна поперечна Т-трубочка та прилеглі до неї мембрани саркоплазматичного ретикулуму. Відстань між Т-трубочками та мембраною саркоплазматичного ретикулуму дорівнює 20 нм; функція тріади – електричний синапс.
При виникненні потенціалу дії в м'язі він поширюється поверхневою плазматичною мембраною, як по безмієліновому нервовому волокну. Потім по Т-системі потенціал дії поширюється вглиб волокна. При цьому через електричний синапс збудження передається на мембрану саркоплазматичного ретикулуму. В результаті підвищується проникність саркоплазматичного ретикулуму для іонів Са2+ і вони виходять у міжфібрилярний простір.
Висновок:провідна система м'язового волокна забезпечує поширення потенціалу дії та виходу Са2+ з саркоплазматичного ретикулуму в міжфібрилярний простір.
Сучасні уявлення про будову скелетного м'яза
Скелетні м'язи складаються з міофібрил, які за допомогою Z-мембрани поділені на окремі саркомери.
Саркомір- це основний скорочувальний елемент кістякових м'язів.
У саркомірі розрізняють:
1) темна частина у центрі саркомера (диск А);
2)у центрі диска А світле простір - Н-мембрана;
3)світлі ділянки саркомера - Диск J.
Диски А та J утворені окремими протофібрилами. А-фібрили товсті з білка міозину, J – тонкі з білка актину. Молекула міозину є тілом з важкого мероміозину і голівку - з легкого мероміозину. На голівці фіксовано молекулу АТФ, яка у спокої заряджена негативно. На підставі головки фіксується молекула ферменту АТФ-ази, також заряджена негативно. Молекули відштовхуються – головка перебуває у розправленому стані. Товсті протофібрили складаються з 3-х білків - тропоміозінова нитка, на яку накручена подвійна спіраль глобулярного актину. Через рівномірні проміжки розташовується білок тропонін - "щит", що закриває А-центр тонкої протофібрили. Тропонін має високу спорідненість до Са2+ Тропонінові центри розташовуються у вигляді спіралі приблизно через кожні 15 нм. За рахунок цих тропонінових комплексів відбувається відкриття А-центру протофібрили та утворюються містки між актиновими та міозиновими нитками.