Медіатором у нервово-м'язовому синапсі є. Речовини, що блокують передачу збудження. Структура та функції нервово-м'язового синапсу
Синапс – це спеціалізована структура, яка забезпечує передачу збудження з однієї збудливої структури на іншу. Термін "синапс" введений Ч. Шеррінгтоном і означає "зведення", "з'єднання", "застібка".
Класифікація синапсів.Синапси можна класифікувати за:
їх місцезнаходження та належність відповідним структурам:
* периферичні (нервово-м'язові, нейро-секреторні, рецепторнонейрональні);
* центральні (аксо-соматичні, аксо-дендрітні, аксо-аксональні, сомато-дендрітні, сомато-соматичні);
знаку їх дії - збуджуючі та гальмівні;
способу передачі сигналів – хімічні, електричні, змішані.
медіатору, за допомогою якого здійснюється передача -холінергічні, адренергічні, серотонінергічні, гліценергічні і т.д.
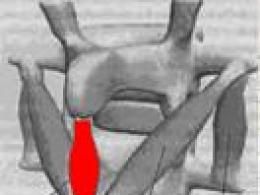
Будова синапсу.Всі синапси мають багато спільного, тому будову синапсу та механізм передачі збудження в ньому можна розглянути на прикладі нервово-м'язового синапсу (рис. 7).
Синапс складається з трьох основних елементів:
* пресинаптичної мембрани - (в нервово-м'язовому синапсі - це потовщена кінцева платівка);
* Постсинаптичної мембрани;
* синаптичної щілини.
Пресинаптична мембрана- це частина мембрани м'язового закінчення області контакту його з м'язовим волокном. Постсинаптична мембрана – частина мембрани м'язового волокна. Частина постсинаптичної мембрани, що розташована навпроти пресинаптичної, називається субсинаптичною мембраною. Особливістю субсинаптичної мембрани є наявність у ній спеціальних рецепторів, чутливих до певного медіатора, та наявність хемозалежних каналів. У постсинаптичній мембрані, за межами субсинаптичної, є потенціалозалежні канали.
Механізм передачі збудження у хімічних збудливих синапсах.У синапсах із хімічною передачею збудження передається за допомогою медіаторів (посередників). Медіатори- це хімічні речовини, які забезпечують передачу збудження у синапсах. Медіатори в залежності від їхньої природи діляться на кілька груп:
* моноаміни(ацетилхолін, дофамін, норадреналін, серотонін та ін);
* амінокислоти (гама-аміномасляна кислота - ГАМК, глутамінова кислота, гліцин та ін);
* нейропептиди(речовина Р, ендорфіни, нейротензин, АКТГ, ангіотензин, вазопресин, соматостатин та ін.).
Медіатор у молекулярному вигляді знаходиться у бульбашках пресинаптичного потовщення (синаптичної бляшки), куди він надходить:
* з навколоядерної області нейрона за допомогою швидкого аксонального транспорту (аксотока);
* за рахунок синтезу медіатора, що протікає в синаптичних терміналях із продуктів його розщеплення;
* за рахунок зворотного захоплення медіатора із синаптичної щілини в незмінному вигляді.
Коли за аксоном до його терміналів приходить збудження, пресинаптична мембрана деполяризується, що супроводжується надходженням іонів кальцію із позаклітинної рідини всередину нервового закінчення. Іони кальцію, що надійшли, активують переміщення синаптичних бульбашок до пресинаптичної мембрани, їх дотик і руйнування (лізис) їх мембран з виходом медіатора в синаптичну щілину. У ній медіатор дифундує до субсинаптичної мембрани, де знаходяться його рецептори. Взаємодія медіатора з рецепторами призводить до відкриття каналів переважно для іонів натрію. Це призводить до деполяризації субсинаптичної мембрани та виникнення так званого збудливого постсинаптичного потенціалу (ВПСП). У нервово-м'язовому синапсі ВПСП називається потенціалом кінцевої платівки (ПКП). Між деполяризованою субсинаптичною мембраною та сусідніми з нею ділянками постсинаптичної мембрани виникають місцеві струми, які деполяризують мембрану. Коли вони деполяризують мембрану до критичного рівня, у постсинаптичній мембрані м'язового волокна виникає потенціал дії, який поширюється мембранами м'язового волокна і викликає його скорочення.
Хімічні гальмівні синапси.Ці синапси за механізмом передачі збудження подібні до синапсів збудливої дії. У гальмівних синапсах медіатор (наприклад, гліцин) взаємодіє з рецепторами субсинаптичної мембрани і відкриває в ній хлорні канали, це призводить до руху іонів хлору концентраційним градієнтом всередину клітини та розвитку гіперполяризації на субсинаптичній мембрані. Виникає так званий гальмівний постсинаптичний потенціал (ТПСП).
Раніше вважали, що кожному медіатору відповідає специфічна реакція постсинаптичної клітини – збудження чи гальмування у тій чи іншій формі. В даний час встановлено, що одному медіатору найчастіше відповідає не один, а кілька різних рецепторів. Наприклад, ацетилхолін у нервово-м'язових синапсах скелетних м'язів діє на Н-холінорецептори (чутливі до нікотину), які відкривають широкі канали для натрію (і калію), що породжує ВПСП (ПКП). У вагосерцевих синапсах той же ацетилхолін діє на М-холінорецептори (чутливі до мускарину), що відкривають селективні канали для іонів калію, тому тут генерується гальмівний постсинаптичний потенціал (ТПСП). Отже, збуджуючий або гальмівний характер дії медіатора визначається властивостями субсинаптичної мембрани (точніше видом рецептора), а не самого медіатора.
Фізіологічні властивості хімічних синапсів.
Синапси з хімічною передачею збудження мають ряд загальних властивостей:
* Порушення через синапс проводиться тільки в одному напрямку (односторонньо). Це зумовлено будовою синапсу: медіатор виділяється лише з пресинаптичного потовщення та взаємодіє з рецепторами субсинаптичної мембрани;
* передача збудження через синапс здійснюється повільніше, ніж по-нервовому волокну - синаптична затримка;
* передача збудження здійснюється за допомогою спеціальних хімічних посередників – медіаторів;
* у синапсах відбувається трансформація ритму збудження;
* синапси мають низьку лабільність;
* синапси мають високу стомлюваність;
* синапси мають високу чутливість до хімічних (у тому числі і фармакологічних) речовин.
Електричні синапси збудливої дії.Окрім синапсів із хімічною передачею збудження переважно у центральній нервовій системі (ЦНС) зустрічаються синапси з електричною передачею. Збудливим електричним синапсам властиві дуже вузька синаптична щілина та дуже низький питомий опір зближених пре- та постсинаптичних мембран, що забезпечує ефективне проходження локальних електричних струмів. Низький опір, як правило, пов'язаний з наявністю поперечних каналів, що перетинають обидві мембрани, тобто що йдуть з клітини в клітину (щілинний контакт). Канали утворюються білковими молекулами (напівмолекулами) кожної з контактуючих мембран, які з'єднуються комплементарно. Ця структура легко прохідна електричного струму.
Схема передачі збудження в електричному синапсі: струм, викликаний пресинаптичним потенціалом дії, подразнює постсинаптичну мембрану, де виникає ВПСП та потенціал дії.
Поперечні канали об'єднують клітини як електрично, а й хімічно, оскільки вони прохідні багатьох низькомолекулярних сполук. Тому збуджуючі електричні синапси з поперечними каналами формуються, як правило, між клітинами одного типу (наприклад, між клітинами серцевого м'яза).
Загальними властивостями збуджувальних електричних синапсів є:
* Швидкодія, (значно перевищує таке в хімічних синапсах);
* Слабкість слідових ефектів при передачі збудження (в результаті цього в них практично неможлива сумація послідовних сигналів);
* Висока надійність - передачі збудження.
Збудливі електричні синапси можуть виникати за сприятливих умов і зникати при несприятливих. Наприклад, при пошкодженні однієї з клітин, що контактують, її електричні синапси з іншими клітинами ліквідуються. Ця властивість називається пластичністю.
Електричні синапси можуть бути з односторонньою та двосторонньою передачею збудження.
Електричний гальмівний синапс. Поряд із електричними синапсами збудливої дії можуть зустрічатися електричні гальмівні синапси. Прикладом такого синапс може служити синапс, який утворює нервове закінчення на вихідному сегменті маутнерівського нейрона у риб. Гальмуючий вплив виникає за рахунок дії струму, спричиненого потенціалом дії пресинаптичної мембрани. Пресинаптичний потенціал викликає значну гіперполяризацію сегмента та гіперполяризуючий струм миттєво гальмує генерацію потенціалу дії у початковому сегменті аксону.
У змішаних синапсахпресинаптичний потенціал дії створює струм, який деполяризує постсинаптичну мембрану типового хімічного синапсу, де пре- та постсинаптичні мембрани нещільно прилягають одна до одної. Таким чином, у цих синапсах хімічна передача служить необхідним механізмом, що підсилює.
ФІЗІОЛОГІЯ ЦЕНТРАЛЬНОЇ НЕРВОВОЇ СИСТЕМИ.
ФУНКЦІЇ ЦЕНТРАЛЬНОЇ НЕРВОВОЇ СИСТЕМИ. Організм людини є складною високоорганізованою системою, що складається з функціонально пов'язаних між собою клітин, тканин, органів та їх систем.
Цей взаємозв'язок (інтеграцію) функцій, їх узгоджене функціонування забезпечує центральна нервова система (ЦНС). ЦНС регулює всі процеси, які у організмі, тому з її допомогою відбуваються найбільш адекватні зміни роботи різних органів, створені задля забезпечення тій чи іншій його діяльності.
ЦНС також здійснює зв'язок організму із зовнішнім середовищем, шляхом аналізу і синтезу різноманітної інформації, що надходить до неї від рецепторів. Вона виконує функції регулятора поведінки, який буде необхідний у конкретних умовах існування. Це забезпечує адекватне пристосування до навколишнього світу. Крім того, з функціями ЦНС пов'язані процеси, що лежать в основі психічної діяльності людини.
МЕТОДИ ДОСЛІДЖЕННЯ ФУНКЦІЙ ЦНС. Інтенсивний розвиток функцій ЦНС зумовив перехід від описових методів вивчення функцій різних відділів мозку до експериментальних методів. Багато методів, що використовуються для вивчення функції ЦНС, застосовують у поєднанні один з одним.
Метод руйнування (екстерпації) різних відділів центральної нервової системи. За допомогою цього можна встановити які функції ЦНС випадають після оперативного втручання і які зберігаються. Даний методичний прийом давно використовується в експериментально-фізіологічних дослідженнях.
Метод перерізання дає можливість вивчити значення діяльності того чи іншого відділу ЦНС впливів, що надходять від інших її відділів. Перерізування проводиться на різних рівнях центральної нервової системи. Повна перерізка, наприклад, спинного мозку або стовбура мозку роз'єднує вищележачі відділи ЦНС від нижчележачих і дозволяє вивчити рефлекторні реакції, що здійснюються нервовими центрами, розташованими нижче місця перерізання. Перерізання та локальне пошкодження окремих нервових центрів проводиться не тільки в умовах експерименту, але й у нейрохірургічній клініці як лікувальні заходи.
Метод подразнення дозволяє вивчити функціональне значення різних утворень ЦНС. При подразненні (хімічному, електричному, механічному тощо) певних структур мозку можна спостерігати виникнення, особливості прояви та характер поширення процесів збудження.
Електроенцефалографія – метод реєстрації сумарної електричної активності різних відділів головного мозку. Вперше запис електричної активності мозку було здійснено В. В. Правдич-Немінським за допомогою електродів, занурених у мозок. Бергер зареєстрував потенціали мозку з поверхні черепа та назвав запис коливань потенціалів мозку електроенцефалограмою (ЕЕГ-ма).
Частота та амплітуда коливань може змінюватися, але в кожний момент часу в ЕЕГ-мі переважають певні ритми, які Бергер назвав альфа-, бета-, тета- та дельта-ритмами. Альфа-ритм характеризується частотою коливань 8-13 Гц, амплітуда 50 мкВ. Цей ритм найкраще виражений у потиличній та тім'яній ділянці кори та реєструється в умовах фізичного та розумового спокою при закритих очах. Якщо очі відкрити, то альфа-ритм змінюється швидшим бета-ритмом. Бета-ритм характеризується частотою коливань 14-50 Гц та амплітудою до 25 мкВ. У деяких людей альфа-ритм відсутня, і тому у спокої реєструється бета-ритм. У зв'язку з цим розрізняють бета-ритм 1 з частотою коливань 16-20 Гц, він характерний для стану спокою і реєструється у лобовій та тім'яній областях. Бета-ритм 2 з частотою 20-50 Гц і характерний для стану інтенсивної діяльності мозку. Тета-ритм є коливаннями з частотою 4-8 Гц і амплітудою 100-150 мкВ. Цей ритм реєструється у скроневій та тім'яній областях при психомоторній активності, при стресі, під час сну, при гіпоксії та легкому наркозі. Дельта-ритм характеризується повільними коливаннями потенціалів із частотою 0,5-3,5 Гц, амплітудою 250-300 мкВ. Цей ритм реєструється під час глибокого сну, при глибокому наркозі, гіпоксії.
ЕЕГ метод використовується у клініці з діагностичною метою. Особливо широке застосування цей метод знайшов у нейрохірургічній клініці визначення локалізації пухлин мозку. У неврологічній клініці цей метод знаходить застосування щодо локалізації епілептичного вогнища, в психіатричній клініці - для діагностики розладів психіки. У хірургічній клініці ЕЕГ використовують для тестування глибини наркозу.
Метод викликаних потенціалів – реєстрація електричної активності певних структур мозку при стимуляції рецепторів, нервів, підкіркових структур. Викликані потенціали (ВП) найчастіше є трифазними коливаннями ЕЕГ-ми, що змінюють один одного: позитивне, негативне, друге (пізніше) позитивне коливання. Однак, вони можуть мати більш складну форму. Розрізняють первинні (ПЗ) і пізні або вторинні (ВО) викликані потенціали. ВП - це фрагмент ЕЕГ-ми, записаний в останній момент стимуляції мозку і має таку ж природу, як і електроенцефалограма.
Метод ВП знаходить застосування в неврології та нейрофізіології. За допомогою ВП можна простежити онтогенетичний розвиток провідних шляхів мозку, провести аналіз локалізації представництва сенсорних функцій, провести аналіз зв'язків між структурами мозку, показати кількість перемикань на шляху розповсюдження збудження тощо.
Мікроелектродний метод застосовується для вивчення фізіології окремого нейрона, його біоелектричної активності як у стані спокою, так і за різних впливів. Для цих цілей використовуються спеціально виготовлені скляні або металеві мікроелектроди діаметр кінчика яких становить 0,5-1,0 мкм або трохи більше. Скляні мікроелектроди являють собою мікропіпетки, заповнені розчином електроліту. Залежно від розташування мікроелектроду розрізняють два способи відведення біоелектричної активності клітин - внутрішньоклітинний та позаклітинний.
Внутрішньоклітинне відведення дозволяє реєструвати та вимірювати:
* мембранний потенціал спокою;
* Постсинаптичні потенціали (ВПСП та ТПСП);
* динаміку переходу місцевого збудження в поширюється;
* потенціал дії та її компоненти.
Позаклітинне відведення дає можливість реєструвати:
* Спайкова активність як окремих нейронів, так і, в основному, їх груп, розташованих навколо електрода.
Для точного визначення становища різних структур мозку і запровадження у яких різних микропредметов (електроди, термопари, піпетки та інших.) широке застосування як і електрофізіологічних дослідженнях, і у нейрохірургічної клініці знайшов стереотаксический метод. Його використання ґрунтується на результатах детальних анатомічних досліджень розташування різних структур головного мозку щодо кісткових орієнтирів черепа. За даними таких досліджень створені спеціальні стереотаксичні атласи як різних видів тварин, так людини. В даний час стереотаксичний метод знаходить широке застосування в нейрохірургічній клініці для таких цілей:
* руйнування структур мозку з метою ліквідації станів гіперкінезу, неприборканого болю, деяких психічних розладів, епілептичних порушень та ін;
* Виявлення патологічних епілептогенних вогнищ;
* введення радіоактивних речовин у пухлини мозку та для руйнування цих пухлин;
* коагуляції аневризм мозкових судин;
* здійснення лікувальних електростимуляцій чи гальмування структур мозку.
БУДОВА ЦНС. Структурно-функціональною одиницею ЦНС є нейрон (нервова клітина). Він складається з тіла (соми) та відростків – численних дендритів та одного аксона. Дендрити зазвичай сильно розгалужуються і утворюють безліч синапсів, з іншими клітинами, що визначає їх провідну роль у сприйнятті нейроном інформації. Аксон починається від тіла клітини аксонним горбком, функцією якого є генерація нервового імпульсу, який за аксоном проводиться до інших клітин. Аксон сильно розгалужується, утворюючи безліч колатералей, терміналі яких утворюють синапси з іншими клітинами. Мембрана аксона в області синапс містить специфічні рецептори, здатні реагувати на різні медіатори або нейромодулятори. Тому процес виділення медіатора пресинаптичними закінченнями може ефективно регулюватися іншими нейронами. Крім того, мембрана закінчень містить велику кількість потенціалозалежних кальцієвих каналів, через які іони кальцію надходять усередину закінчення при його збудженні.
У більшості центральних нейронів ПД виникає в області мембрани аксонного пагорба, збудливість якої вдвічі вища за інші ділянки і звідси збудження поширюється по аксону та тілу клітини. Такий спосіб збудження нейрона важливий для здійснення його інтегративної функції, тобто здатності підсумовувати впливи, що надходять на нейрон з різних синаптичних шляхів. Ступінь збудливості різних ділянок нейрона неоднакова, вона найвища в ділянці аксонного горбка, в ділянці тіла нейрона вона значно нижча, і найнижча у дендритів.
Крім нейронів у ЦНС є гліальні клітини, що займають половину об'єму мозку. Периферичні аксони також оточені оболонкою із гліальних клітин-іванівських клітин. Нейрони та гліальні клітини розділені міжклітинними щілинами, які повідомляються одна з одною та утворюють заповнений рідиною міжклітинний простір нейронів та глії. Через цей простір відбувається обмін речовинами між нервовими та гліальними клітинами. Функції клітин глії різноманітні:
* вони є для нейронів опорним, захисним та трофічним апаратом, підтримують певну концентрацію іонів калію та кальцію в міжклітинному просторі;
* активно поглинають нейромедіатори, обмежуючи таким чином їх час
дії та інші функції.
АКСОННИЙ ТРАНСПОРТ. Аксони, крім функції проведення збудження, є каналами транспорту різних речовин. Білки та медіатор, синтезовані в тілі клітини, органели та інші речовини можуть переміщатися аксоном до його закінчення. Це переміщення речовин отримало назву аксонного транспорту. Існує два його види - швидкий та повільний аксонний транспорт.
ШВИДКИЙ АКСОННИЙ ТРАНСПОРТ - це транспорт везикул, мітохондрій та деяких білкових частинок від тіла клітини до закінчення аксона (антероградний транспорт) зі швидкістю 250-400 мм/сут. Він здійснюється спеціальним транспортним механізмом - за допомогою мікротрубочок та нейрофіламентів і подібний до механізму м'язового скорочення.
Швидкий аксонний транспорт від терміналів аксона до тіла клітини, або ретроградний, переміщає лізосоми, везикули, що виникають в закінченнях аксона в ході піноцитозу, наприклад, ацетилхолінестерази, деяких вірусів, токсинів та ін. зі швидкістю 220 мм/добу. Швидкість швидкого антероградного та ретроградного транспорту не залежить від типу та діаметра аксона.
ПОВІЛЬНИЙ АКСОННИЙ ТРАНСПОРТ забезпечує переміщення зі швидкістю 1-4 мм/сут. білків та структур цитоплазми (мікротрубочок, нейрофіламентів, РНК, транспортних та канальних мембранних білків тощо) у дистальному напрямку за рахунок інтенсивності синтетичних процесів у перикаріоні. Повільний аксонний транспорт має особливе значення у процесах зростання та регенерації відростків нейрона.
РОЗВИТОК РЕФЛЕКТОРНОЇ ТЕОРІЇ.
Основним механізмом діяльності ЦНС є рефлекс. Рефлекс - це реакція організму у відповідь на дії подразника, здійснювана за участю ЦНС і спрямована на досягнення корисного результату.
Рефлекс у перекладі з латинської означає "відображення"-. Вперше термін "віддзеркалення" або "рефлектування" був застосований Р. Декартом (1595-1650) для характеристики реакцій організму у відповідь на подразнення органів чуття. Він першим висловив думку, що всі прояви эффекторной активності організму викликаються цілком реальними фізичними чинниками. Після Р. Декарта уявлення про рефлекс було розвинене чеським дослідником Г. Прохазькою, який розвинув вчення про відбивні дії. У цей час вже було зазначено, що у спинальних тварин рухи наступають у відповідь на подразнення певних ділянок шкіри, а руйнування спинного мозку веде до їхнього зникнення.
Подальший розвиток рефлекторної теорії пов'язаний з ім'ям І. М. Сєченова. У книзі "Рефлекси головного мозку" він стверджував, що всі акти несвідомого та свідомого життя за природою походження є рефлексами. Це була геніальна спроба запровадити фізіологічний аналіз у психічні процеси. Але на той час не існувало методів об'єктивної оцінки діяльності мозку, які могли б підтвердити цю пропозицію І. М. Сєченова. Такий об'єктивний метод був розроблений І. П. Павловим - метод умовних рефлексів, за допомогою якого він довів, що найвища нервова діяльність організму, як і нижча, є рефлекторною.
Структурною основою рефлексу, його матеріальним субстратом (морфологічною основою) є рефлекторна дуга - сукупність морфологічних структур, що забезпечує здійснення рефлексу (шлях, яким відбувається збудження під час здійснення рефлексу).
В основі сучасного уявлення про рефлекторну діяльність лежить поняття корисного пристосувального результату, заради якого відбувається будь-який рефлекс. Інформація про досягнення корисного пристосувального результату надходить до ЦНС за ланкою зворотного зв'язку у вигляді зворотної аферентації, яка є обов'язковим компонентом рефлекторної діяльності. Принцип зворотної аферентації запроваджено рефлекторну теорію П. До. Анохиным. Таким чином, за сучасними уявленнями структурною основою рефлексу є не рефлекторна дуга, а рефлекторне кільце, що складається з наступних компонентів (ланок):
* рецептор;
* аферентний нервовий шлях;
* нервовий центр;
* Еферентний нервовий шлях;
* Робочий орган (ефектор);
* Зворотна аферентація (рис. 8).
Аналіз структурної основи рефлексу проводиться шляхом послідовного вимкнення окремих ланок рефлекторного кільця (рецептора, аферентного та еферентного шляху, нервового центру). При вимиканні будь-якої ланки рефлекторного кільця рефлекс зникає. Отже, реалізації рефлексу необхідна цілісність всіх ланок його морфологічної основи.
Клітини ЦНС мають численні зв'язки один з одним, тому нервова система людини може бути представлена як система нейронних ланцюгів (нейронних мереж), що передають збудження та формують гальмування. У цій нервовій мережі збудження може поширюватися від одного нейрона на багато інших нейронів. Процес поширення збудження від одного нейрона на багато інших нейронів отримав назву іррадіації збудження або дивергентного принципупоширення збудження.
Розрізняють два види іррадіації збудження:
* Спрямована або системна іррадіація коли збудження поширюється по певній системі нейронів і формує координовану пристосувальну діяльність організму;
* Безсистемна або дифузна (ненаправлена) іррадіація, хаотичне поширення збудження, при якому координована діяльність неможлива (рис. 9).
У ЦНС одного нейрону можуть сходитися збудження від різних джерел. Ця здатність збуджень сходитися до тих самих проміжних і кінцевих нейронів отримала назву конвергенції збуджень(Рис. 9).
Пресинаптична мембрана нервово-м'язового синапсу є частиною мембрани пресинаптичного закінчення аксона мотонейрону, що обмежує синаптичну щілину. Через неї здійснюється виділення (екзоїтоз) медіатора в синаптичну щілину. Медіатор пресинаптичного закінчення міститься в синаптичних пухирцях діаметром 40 нм. Вони утворюються в комплексі Гольджі, за допомогою швидкого прямого аксонного транспорту доставляються в пресинаптичне закінчення і там заповнюються медіатором і АТФ. У нервово-м'язовому синапсі медіатором є ацетилхолін, який утворюється з ацетилкоензиму А та холіну під дією ферменту холінацетилтрансферази. Везикули розташовані переважно поблизу періодичних потовщень пресинаптичної мембрани, які називають активними зонами. У неактивному синапсі везикули за допомогою білка синапсину пов'язані з білками цитоскелета, що забезпечує їм іммобілізацію та резервування. Важливими структурами пресинаптичного закінчення є мітохондрії, що здійснюють енергетичне забезпечення процесу синаптичної передачі, цистерни гладкої ЕПС, що містять депонований іон Са, мікротрубочки та мікрофіламенти, що беруть участь у внутрішньоклітинному пересуванні везикул.
Синаптична щілина в нервово-м'язовому синапсі має ширину в середньому 50 нм. Вона містить міжклітинну рідину та мукополісахаридну щільну речовину у вигляді смужок, містків, яка називається базальною мембраною та містить ацетилхолінестеразу.
Постсинаптична мембрана містить рецептори, здатні пов'язувати молекули медіатора. Її особливістю є наявність дрібних складок, які утворюють кишені, що відкриваються у синаптичну щілину.
Таким чином, основними етапами передачі збудження в нервово-м'язовому синапсі є:
1) збудження мотонейрону, поширення потенціалу на пресинаптичну мембрану;
2) підвищення проникності пресинаптичної мембрани для іонів кальцію, струм кальцію в клітину, підвищення концентрації кальцію в пресинаптичному закінченні;
3) злиття синаптичних бульбашок з пресинаптичною мембраною в активній зоні, екзоцитоз, надходження медіатора до синаптичної щілини;
4) дифузія ацетилхоліну до постсинаптичної мембрани, приєднання його до N-холінорецепторів, відкриття хемозалежних іонних каналів;
5) переважний іонний струм натрію через хемозалежні канали, утворення надпорогового потенціалу кінцевої пластинки;
6) виникнення потенціалів на м'язової мембрані;
7) ферментативне розщеплення ацетилхоліну, повернення продуктів розщеплення на закінчення нейрона, синтез нових порцій медіатора.
Міорелаксанти-лікарські засоби, що знижують тонус скелетної мускулатури зі зменшенням рухової активності аж до повного знерухомлення.
Механізм дії – блокада Н-холінорецепторів у синапсах припиняє подачу нервового імпульсу до скелетних м'язів, і м'язи перестають скорочуватися. Розслаблення йде знизу нагору, від кінчиків пальців ніг до мімічних м'язів. Останньою розслаблюється діафрагма. Відновлення провідності йде у зворотному порядку.
За особливостями взаємодії з рецепторами міорелаксанти поділяються на дві групи:
Деполяризуючі міорелаксанти - при контакті з рецепторами викликають стійку деполяризацію мембрани синапсу, що супроводжується короткочасним хаотичним скороченням м'язових волокон (міофасцикуляції), що переходить у міорелаксацію. При стійкій деполяризації нервово-м'язова передача припиняється. Міорелаксація нетривала, відбувається за рахунок утримання відкритими мембранних каналів та неможливості реполяризації. Метаболізуються псевдохолінестеразою, виводяться нирками.
Недеполяризуючі міорелаксанти - блокують рецептори та мембранні канали без їх відкриття, не викликаючи деполяризації
20в.Фізіологічні властивості та особливості гладких м'язів
Основні функції м'язової тканини:
1.рухова – забезпечення руху
2.статична – забезпечення фіксації, зокрема й у певній позі
3.рецепторна – у м'язах є рецептори, що дозволяють сприймати власні рухи
4. депонуюча – у м'язах запасаються вода та деякі поживні речовини.
Фізіологічні властивості:
Короткість. Скорочення гладкого м'яза визначається особливим характером поширення збудження. Група клітин, що взаємодіють за допомогою нексусів і своїх електричних полів, формує пучок, який є структурно-функціональною одиницею гладкого м'яза і скорочується як єдине ціле. велика тривалість скорочення; Час скорочення гладком'язового волокна в кілька сотень разів більший, ніж поперечно. Завдяки цьому гладкі м'язи пристосовані до тривалого скорочення без великих витрат енергії та повільно втомлюються;
Спонтанна міогенна активність. На відміну від скелетних м'язів гладкі м'язи шлунка, кишечника, матки, сечоводів, кровоносних судин та інших внутрішніх органів розвивають спонтанно-тетаноподібні скорочення. Від цих клітин ПД поширюється зі швидкістю приблизно 0.1 м/с через Нексус на сусідні волокна та охоплює весь м'яз. Наприклад, перистальтичні скорочення шлунка виникають із частотою 3 рази за 1 хв, сегментарні та маятникові рухи в товстому кишечнику - з частотою -20 разів за 1 хв.
Пластичність – здатність зберігати довжину, набуту при розтягуванні, без зміни напруги. Ця властивість має дуже велике значення для нормальної діяльності внутрішніх органів, наприклад, сечового міхура.
висока чутливість до фізіологічно активних речовин, зокрема до медіаторів вегетативної нервової системи – ацетилхоліну, а також – серотоніну, брадикініну, простагландинів. Зазначені біологічно активні речовини можуть збуджувати, так і гальмувати гладком'язові волокна. Це залежить від того, який процес – деполяризацію, чи гіперполяризацію викликає цю речовину на мембрані клітини. Так, наприклад, ацетилхолін викликає скорочення гладких м'язів більшості органів, але сприяє розслабленню стінок судин деяких органів. Характер відповіді гладких м'язів на дію фізіологічно активної речовини залежить від того, іонні канали вона відкриває, у свою чергу, детерміновано специфікою мембранних рецепторів.
Збудливість. Потенціал спокою 60-70 мВ. Для міоцитів, що мають спонтанну електричну активність 30-60. Позитивний пік ПД менше, ніж у поперечносмугастих менше, ніж у поперечносмугастих м'язових волокнах і досягає 10-15 мВ. Тривалість ГД коливається від 25 мс до 1 с. В процесі формування потенціалу спокою грають роль не тільки іони К+, але і Са) Взаємодія актину та міозину в гладком'язових волокнах також активується іонами Са2, але вони потрапляють у клітини не з сарко-ретикулуму, а транспортуються туди з міжклітинного середовища. Деполяризація мембрани обумовлена відкриттям кальцієвих каналів та дифузією іонів кальцію в клітину.
Провідність. Проведення збудження по гладком'язовому міоциту безперервне. Однак ізольовано окремі гладком'язові клітини не збуджуються та не скорочуються. Взаємодія між окремими міоцитами здійснюється завдяки щілиноподібним контактам, що мають низький електричний опір. Завдяки цьому електричне поле клітини 1 забезпечує збудження інший. Швидкість поширення ПД у межах пучка становить 5-10 м/с.
Автоматія властива клітин-водія ритму (пейсмекерам). У її основі лежить повільна деполяризація, що спонтанно виникає, – при досягненні критичного потенціалу виникає ПД. Ця деполяризація переважно зумовлена дифузією іонів кальцію у клітину.
2.Рефлекторна дуга- Сукупність структур за допомогою якої здійснюється рефлекс. Схематично рефлекторну дугу можна зобразити із 5 ланок.
1. Сприймаюча ланка (рецептор) забезпечує сприйняття змін зовнішнього та внутрішнього середовища організму за допомогою трансформації енергії подразнення у рецепторний потенціал.
Рефлексогенна зона – сукупність рецепторів, подразнення яких викликає рефлекс. При будь-якому подразненні виникають рецепторні потенціали, що забезпечують посилку н. у ЦНС за допомогою 2 ланки.
2. Аферентна ланка
Роль: передача сигналу в цнс до третьої ланки рефлекторної дуги. Для соматичної нервової системи - це аферентний нейрон з його відростками, його тіло розташоване в спинномозкових гангліях або гангліях черепних нервів. Імпульс по дендриту аферентного нейрона, потім за аксоном, далі в цнс.
3. Керуюча ланка - сукупність центральних (для ВНС і ПНР) нейронів, що формують реакцію у відповідь організму.
4. Еферентна ланка – аксон ефекторного нейрона (для соматичної н.с. мотонейрону).
5. Еффектор (робочий орган) ефекторний нейрон соматичної н.с. є мотонейрон
Класифікація рефлексів
1. За умовами появи рефлексів в онтогенезі
a) Вроджені (безумовні)
b) Придбані
Вроджені можуть бути соматичними (за допомогою сомат. н.с., як ефектор – скелетна мускулатура) і вегетативними (за допомогою вегет. н.с.)
2. За біологічним значенням
a) Гомеастатичні (регуляція функцій внутр. органів; робота серця; секреція та моторика ШКТ – харчові рефлекси.)
b) Захисні (оборонні)
c) Статеві
d) орієнтовний рефлекс.
3. Залежно від кількості синапсів
У центральній частині рефлекторної дуги розрізняють.
а) моносинаптичні (рефлекс на розтяг чотириголовного м'яза - колінний розгинальний рефлекс, при ударі по сухожиллю
б) полісинаптичні (бере участь кілька послідовно включених нейронів ЦНС)
4. за рецепторами, подразнення яких викликає реакцію у відповідь.
а) екстероцептивні
б) інтероцептивні
в) пропріоцептивні (використовуються у клінічній практиці для оцінки стану збудливості ЦНС та для діагностики.
5. з локалізації рефлекторної дуги
А) центральні (дуга через ЦНС)
б) периферичні ревлекси (дуга замикається поза ЦНС)
в) стосовно фізіологічних систем
|
ОЗНАКИ |
Вегетативна |
СОМАТИЧНА |
|
Органи-мішені |
Гладкі м'язи, міокард, залози, жирова тканина, органи імунітету |
Скелетні м'язи |
|
Паравертебральні, Превертебральні та Органні |
Локалізовано в ЦНС |
|
|
Число еферентних нейронів | ||
|
Ефект стимуляції |
Збудливий або Переважний |
Збудливий |
|
Типи нервових волокон |
Тонкі мієлінові або немієлінові, повільні |
мієлінові швидкі |
Сінапс(грец. synapsis- з'єднання) - це спеціалізована структура, що забезпечує передачу сигналу від клітини до клітини. Через синапс реалізується дія багатьох фармакологічних препаратів.
Структурно-функціональна організація.Кожен синапс має пре- І постсинаптичну мембрануі синаптичну щілину(Рис. 17).
Рис. 17. Нервово-м'язовий синапс кістякового м'яза: 1 – гілка аксона; 2 – пресинаптичне закінчення аксона; 3 – мітохондрія; 4 – синаптичні бульбашки, що містять ацетилхолін; 5 – синаптична щілина; 6 – молекули медіатора у синаптичній щілині; 7 – постсинаптична мембрана м'язового волокна з N-холінорецепторами
Пресинаптична мембрананервово-м'язового синапсу є частиною мембрани пресинаптичного закінчення аксона мотонейрона. Через неї здійснюється викид (екзоцитоз) медіатора(Лат. mediator- посередник) у синаптичну щілину. У нервово-м'язовому синапсі медіатором є ацетилхолін. Медіатор пресинаптичного закінчення міститься у синаптичних пухирцях (везикулах), діаметр яких становить близько 40 нм. Вони утворюються в комплексі Гольджі, за допомогою швидкого аксонного транспорту доставляються в пресинаптичне закінчення, де заповнюються медіатором та АТФ. У пресинаптичному закінченні міститься кілька тисяч везикул, у кожній з яких є від 1 до 10 тис. молекул хімічної речовини.
Постсинаптична мембрана (кінцева платівкав нервово-м'язовому синапсі) - це частина клітинної мембрани иннервируемой м'язової клітини, що містить рецептори, здатні пов'язувати молекули ацетилхоліну. Особливість цієї мембрани: безліч дрібних складок, що збільшують її площу і кількість рецепторів на ній до 10-20 млн в одному синапсі.
Синаптична щілинау нервово-м'язовому синапсі має ширину в середньому 50 нм. Вона містить міжклітинну рідину, ацетилхолінестеразу і мукополісахаридну щільну речовину у вигляді смужок, містків, у сукупності утворюють базальну мембрану, що з'єднує пре-і постсинаптичну мембрани.
Механізми синаптичної передачівключають триосновних етапу(Рис. 18).
Рис. 18. Механізм проведення імпульсу через хімічний синапс: 1-8 – етапи процесу (Чеснокова, 2007)
Перший етап- процес викиду медіаторав синаптичну щілину, який запускається за допомогою ПД пресинаптичного закінчення. Деполяризація його мембрани веде до відкриття потенціал керованих Са-каналів. Са 2+ входить у нервове закінчення згідно з електрохімічним градієнтом. Частина медіатора у пресинаптичному закінченні локалізується на пресинаптичній мембрані зсередини. Са 2+ активує екзоцитозний апарат пресинапсу, що є сукупністю білків (синапсин, спектрин та ін), пресинаптичного закінчення, активація яких забезпечує викид ацетилхоліну за допомогою екзоцитозу в синаптичну щілину. Кількість ацетилхоліну, що вивільняється, з пресинаптичного закінчення пропорційно в четвертому ступені кількості надійшов туди Са 2+ . На один ПД із пресинаптичного закінчення нервово-м'язового синапсу викидається 200-300 квантів (везикул) медіатора.
Другий етап - дифузія ацетилхолінупротягом 0,1-0,2 мс до постсинаптичної мембрани та дія його на N-холінорецептори (стимулюються також нікотином, внаслідок чого і отримали свою назву). Видалення ацетилхоліну з синаптичної щілини здійснюється шляхом руйнування його під дією ацетилхолінестерази, розташованої в базальній мембрані синаптичної щілини, протягом кількох десятих часток мілісекунди. Близько 60% холіну захоплюється назад пресинаптичним закінченням, що робить синтез медіатора більш економічним, частина ацетилхоліну розсіюється . У проміжках між ПД із пресинаптичного закінчення відбувається спонтанне виділення 1-2 квантів медіатора в синаптичну щілину протягом 1 с, формуючи так звані мініатюрні потенціали(0,4-0,8 мВ). Вони підтримують високу збудливість клітини, що іннервується, в умовах функціонального спокою і виконують трофічну роль, а в ЦНС - сприяють підтримці тонусу її центрів.
Третій етап - взаємодія ацетилхолінуз N-холінорецепторами постсинаптичної мембрани, внаслідок чого відкриваються іонні канали на 1 мс і, внаслідок переважання входу N+ у клітину, відбувається деполяризація постсинаптичедкої мембрани (кінцевої пластинки). Цю деполяризацію в нервово-м'язовому синапсі називають потенціалом кінцевої платівки(ПКП) (рис. 19).
Особливістю нервово-м'язового синапсу скелетного м'язового волокна і те, що з одиночної його активації формується ПКП великий амплітуди (30-40 мВ), електричне полі якого викликає генерацію ПД на мембрані м'язового волокна поблизу синапса. Велика амплітуда ПКП обумовлена тим, що нервові закінчення поділяються на численні гілочки, кожна з яких викидає медіатор.

Рис. 19. Потенціал кінцевої платівки (Шмідт, 1985): КП- Критичний потенціал; ПД -потенціал дії; А- ПКП у нормальному м'язі; Б– ослаблений ПКП у курарезованому м'язі; стрілкамивказано момент нанесення стимулу
Характеристика проведення збудження у хімічних синапсах. Одностороннє проведення збудженнявід нервового волокна до нервової або ефекторної клітини, оскільки пресинаптичне закінчення чутливе лише до нервового імпульсу, а постсинаптична мембрана – до медіатора.
Неізольоване- Порушення поряд розташованих постсинаптичних мембран підсумовується.
Синаптична затримкау передачі сигналу до іншої клітини (в нервово-м'язовому синапсі 0,5-1,0 мс), що пов'язано з вивільненням медіатора з нервового закінчення його дифузією до постсинаптичної мембрани і виникненням постсинаптичних потенціалів, здатних викликати ПД.
Декрементність (згасання) збудження в хімічних синапсах при недостатньому виділенні медіатора із пресинаптичних закінчень у синаптичні щілини.
Низька лабільність(У нервово-м'язовому синапсі становить 100 Гц), яка в 4 - 8 разів нижча за лабільність нервового волокна. Це пояснюється синаптичною затримкою.
Провідність нервово-м'язового синапсу (як і хімічних синапсів ЦНС) пригнічується або, навпаки, стимулюється різними речовинами.
Наприклад, кураре та курареподібні речовини (диплацин, тубокурарин) оборотно зв'язуються з N-холінорецепторами постсинаптичної мембрани, блокують дію на неї ацетилхоліну та передачу в синапсі. Навпаки, деякі фармакологічні препарати, наприклад прозерин, пригнічують активність ацетилхолінестерази, сприяючи помірному накопиченню ацетилхоліну та полегшенню синаптичної передачі, що використовується в лікувальній практиці.
Стомлюваність(Синаптична депресія) - погіршення провідності аж до повної блокади проведення збудження при тривалому функціонуванні синапсу (головна причина - виснаження медіаторау пресинаптичному закінченні).
Запитання для самоконтролю
1. Який механізм поширення збудження по нервовому волокну? Яка роль перехоплень Ранв'є у проведенні збудження по мієлінізованому нервовому волокну?
2.У чому перевага стрибкоподібного (сальтаторного) поширення збудження над безперервним його проведенням вздовж мембрани волокна?
3.У чому фізіологічне значення ізольованого проведення збудження з нервового волокна?
4. Які нервові волокна (аферентні чи еферентні, вегетативні чи соматичні) відносяться до групи А? Яка швидкість проведення збудження за ними?
5. Які нервові волокна (аферентні чи еферентні, вегетативні чи соматичні) відносяться до групи В? Яка швидкість проведення за ними?
6. Які нервові волокна (аферентні чи еферентні, вегетативні чи соматичні) відносяться до групи С? Яка швидкість проведення збудження за ними?
7. Перерахуйте структури нервово-м'язового синапсу (скелетний м'яз). Що називають кінцевою платівкою?
8. Перерахуйте послідовність процесів, що ведуть до звільнення медіатора з пресинаптичної мембрани в синаптичну щілину під час передачі збудження в синапсі.
9. Локальним потенціалом або збудженням, що поширюється, є потенціал кінцевої платівки?
10.Що таке мініатюрні потенціали кінцевої платівки, який механізм їх виникнення?
11.У чому полягає трофічний вплив нерва на м'яз, що здійснюється через нервово-м'язовий синапс?
12. Які речовини є медіаторами в нервово-м'язових синапсах гладкого і поперечного м'язів?
13. Що таке сенсорний рецептор?
14.На які дві групи діляться сенсорні рецептори за швидкістю адаптації? Назвіть рецептори, які стосуються кожної з них.
15. Що розуміють під первинними та вторинними рецепторами?
16. Перерахуйте основні властивості рецепторів.
17. Що називають адаптацією рецепторів? Як змінюється частота імпульсів аферентному нервовому волокні при адаптації рецептора?
18.Назвіть локальні потенціали, що виникають при збудженні первинних та вторинних рецепторів.
19. Рецепторний потенціал, де він виникає, яке його значення?
20.Генераторний потенціал, де він виникає, яке його значення?
21.Де виникає потенціал дії при збудженні первинного сенсорного рецептора?
22. Де виникає потенціал дії під час порушення вторинного сенсорного рецептора?
Фізіологія м'язів
1.3.1. Структурнофункціональна характеристика скелетного м'яза
М'язиподіляють на поперечносмугасті (скелетнаі серцева) та гладкі(судини та внутрішні органи, крім серця).
Скелетний м'язскладається з м'язових волокон, ізольованих у структурному та функціональному відношенні один від одного, які являють собою витягнуті багатоядерні клітини. Товщина волокна становить 10-100 мкм, а його довжина варіює в межах від кількох міліметрів до кількох сантиметрів. Кількість м'язових волокон, встановившись постійним на 4-5-му місяці постнатального онтогенезу, надалі не змінюється; з віком змінюються (збільшуються) лише їх довжина та діаметр.
Призначення основних структурних елементів.Характеристика основних елементів м'язового волокна. Від клітинної мембрани м'язового волокна (сарколеми) вглиб відходять численні поперечні інвагінації ( Т-трубочки), які забезпечують її взаємодію з саркоплазматичним ретикулулом ( СПР) (рис. 20).

Рис. 20. Взаємини клітинної мембрани (1), поперечних трубочок (2), бічних цистерн (3) і поздовжніх трубочок (4) саркоплпзматичного ретикулуму, скорочувальних білків (5): А – у стані спокою; Б - при скороченні м'язового волокна; точками позначені іони Ca 2+
СПРявляє собою систему зв'язаних один з одним цистерн і відходять від них у поздовжньому напрямку канальців, розташованих між міофібрил. Термінальні (кінцеві) цистерни СПР примикають до Т-трубочок, формуючи так звані тріади. У цистернах міститься Са 2+ , що відіграє у м'язовому скороченні. У саркоплазмі є внутрішньоклітинні елементи : ядра, мітохондрії, білки (у тому числі міоглобін), крапельки жиру, гранули глікогену, фосфатсодержащіе речовини, різні малі молекули та електроліти.
Міоібрили- субодиниці м'язового волокна. В одному м'язовому волокні може налічуватися понад 2 тис. міофібрил, їх діаметр 1-2 мкм. В одиночній міофібрилі міститься 2-2,5 тис. протофібрил- Паралельно розташованих ниток білка ( тонкі - актин, товсті - міозин). Актинові нитки складаються з двох субодиниць, скручених у вигляді спіралі. До складу тонких ниток входять також регуляторні білки. тропоміозин та тропонін(Рис. 21).
Рис. 21. Взаємне розташування структурних елементів міофібрил при їх розслабленні (А, Б) та скороченні (В)
Ці білки в незбудженому м'язі перешкоджають взаємозв'язку актину та міозину, тому м'яз у спокої перебуває в розслабленому стані. Міофібрили включають послідовно з'єднані блоки - саркомери(Б), відокремлені один від друга Z-смужками.Саркомір (довжина 2-Змкм) є скоротливою одиницею м'язового волокна; при довжині 5см воно включає близько 20 тис. послідовно з'єднаних саркомерів. Міофібрили окремого м'язового волокна пов'язані таким чином, що розташування саркомерів збігається, і це створює картину поперечної смугастість волокна при спостереженні у світловому мікроскопі (рис. 22).
Рис. 22. Саркомір міоциту скелетного м'яза (A. Vander, J. Sherman, D. Luciano, 2004)
Елементи саркоміру(Див. рис. 21). Міозинові протофібрили утворюють темну частину саркомера - А-диск(Анізотропний, він сильно поляризує біле світло). Світлішу ділянку в центрі А-диску називають Н-зоною. Світлу ділянку саркомера між двома А-дисками називають 1-диском(Ізотропне, майже не поляризує світло). Він утворений актиновими протофібрилами, що йдуть в обидва боки від Z-смужок. Кожен саркомер має два набори тонких ниток, прикріплених до Z-смужок, та один комплект товстих ниток, зосереджених в А-диску. У розслабленому м'язі кінці товстих і тонких філаментів різною мірою перекривають один одного на межі між А- та 1-дисками.
Класифікація м'язових волокон:
За структурно-функціональними властивостямита кольору виділяють дві основні групи м'язових волокон: швидкі та повільні.
Білі (швидкі)м'язові волокна містять більше міофібрил і менше - мітохондрій, міоглобіну та жирів, але більше глікогену та гліколітичних ферментів; ці волокна називають гліколітичні. Капілярна мережа, що оточує ці волокна, щодо рідкісна. Швидкість робочого циклу у даних волокон приблизно в 4 рази більше, ніж у повільних, що пояснюється більш високою АТФазною активністю швидких волокон, але вони мають малу витривалість. У білих м'язових волокон число ниток актину та міозину більше, ніж у червоних, тому вони товщі і сила їх скорочення більше, ніж у червоних волокон.
Червоні м'язові волокнамістять багато мітохондрій, міоглобінужирних кислот. Ці волокна оточені густою мережею кровоносних капілярів, мають менший діаметр. Мітохондрії забезпечують високий рівень окисного фосфорилювання, тому дані волокна називають оксидативними. Червоні м'язові волокна поділяютьсяна дві підгрупи: швидкі та повільні. Повільні волокна можутьвиконувати роботу протягом щодо тривалого часу; стомлення у них розвивається повільніше. Вони більш пристосовані до тонічних скорочень. Червоні швидківолокна за швидкістю стомлення займають проміжне положення між білими та червоними повільними. Швидкість їх скорочення близька до швидкості скорочення білих волокон, що також пояснюється високою активністю АТФаз міозину червоних швидких волокон.
Також є незначна кількість справжніх тонічних м'язових волокон; на них локалізується по 7-10 синапсів, що належать, як правило, кільком мотонейронам, наприклад, в окорухових м'язах, м'язах середнього вуха. ПКП цих м'язових волокон не викликають генерації ПД у них, а безпосередньо запускають м'язове скорочення.
Група м'язових волокон, рухову (нейромоторну) одиницю.У м'язах, що здійснюють швидкі та точні рухи, наприклад у окорухових, нейромоторні одиниці складаються з 3-5 м'язових волокон. У м'язах, що здійснюють менш точні рухи (наприклад, м'язи тулуба та кінцівок), рухові одиниці включають сотні та тисячі м'язових волокон. Велика рухова одиниця, порівняно з малою, включає великий мотонейрон із відносно товстим аксоном, який утворює велику кількість кінцевих гілочок у м'язі і, отже, іннервує велику кількість м'язових волокон. Усі м'язові волокна однієї рухової одиниці, незалежно від кількості, відносяться до одного типу. Усі скелетні м'язи за складом є змішаними, тобто. утворені червоними та білими м'язовими волокнами.
Специфічною властивістю всіх м'язів є скоротливість- Здатність скорочуватися, тобто. коротшати або розвивати напругу. Реалізація цієї здатності здійснюється за допомогою збудження та його проведення по м'язовому волокну (Властивості відповідно до збудливості та провідності).
Скелетні м'язи не мають автоматії, керуються організмом довільно імпульсацією з ЦНС, тому їх називають також довільними. Гладкі м'язи за власним бажанням не скорочуються, тому їх називають також мимовільними,але вони мають автоматією.
Функції скелетного м'яза:
Забезпечення рухової активності організму- пошук і добування води та їжі, її захоплення, жування, ковтання, оборонні реакції, трудова діяльність - фізична та творча робота художника, письменника, вченого, композитора зрештою виявляється у русі: малювання, лист, гра на музичному інструменті тощо. п.
Забезпечення дихання(Рухів грудної клітини та діафрагми).
Комунікативна функція(Усна та письмова мова, міміка та жести).
Участьу процесах терморегуляціїорганізму за допомогою зміни інтенсивності скорочувального термогенезу
©2015-2019 сайт
Усі права належати їх авторам. Цей сайт не претендує на авторства, а надає безкоштовне використання.
Дата створення сторінки: 2016-04-15
Нервове волокно безпосередньо не контактує з м'язовим. Між ними дуже вузька щілина (20-50 нм). Нервове закінчення, частина м'язового волокна, до якої прилягає це закінчення, і щілину між ними становлять нервово-м'язовий синапс. Нервове волокно закінчується синаптичною бляшкою, в якій містяться Мтохондрії та значна кількість (близько 300 тис.) бульбашок, де концентрується медіатор – речовина, за допомогою якої збудження поширюється з нерва на м'яз. У нервово-м'язовому синапсі цим медіатором є ацетилхолін. Синтез ацетилхоліну відбувається у синаптичній бляшці і потребує енергетичних витрат. Частина бляшки, що прилягає до щілини, називається пресинаптичною мембраною. З іншого боку щілини розташована постсинаптична мембрана, що належить м'язовому волокну. Частина цієї мембрани, яка безпосередньо прилягає до бляшки, називається кінцевою платівкою.
До складу постсинаптичної мембрани входять рецепторні білки (молекулярна маса – близько 275 000), які реагують на ацетилхолін та називаються холінорецепторами. Вони реагують також на нікотин, звідси їхня назва – Н-холінорецептори. Реакція ацетилхоліну з Н-холінорецепторами наводить конформаційні зміни молекули рецептора. Це впливає на розташований поруч хемочутливий іонний канал, який може пропускати Na +, К +, Са2 +. Білкові структури цих каналів мають негативний заряд і тому аніони через них не проходять.
Передача інформації через нервово-м'язовий синапс відбувається у такій послідовності:
1. Нервовим волокном у бляшки приходить ПД.
2. Внаслідок дії ПД на мембрану нервового закінчення відкриваються Са2+-канали і ці іони вступають у бляшку.
3. Підвищення у бляшці концентрації Са2+ призводить (за участю кальмодуліну) до виходу медіатора з бульбашок у синаптичну щілину. Внаслідок дії одиничного ПД медіатор виходить приблизно із 300 бульбашок.
4. Ацетилхолін дифундує через щілину.
5. Ацетилхолін реагує з Н-холінорецепторами.
6. Відкриваються «ворота» хемочутливих каналів і за концентраційним градієнтом Na+ переходить у м'язове волокно, а К+ – назовні.
7. Відбувається деполяризація кінцевої платівки та розвивається її потенціал (ПКП). ПКП-один із видів місцевого збудження. Чим більше виділяється ацетилхоліну, то більшою мірою виражений цей потенціал.
8. Коли ЛКП досягає критичної величини (ЕКС), деполяризується колосинаптична електрозбудлива мембрана та розвивається
ПД м'язового волокна, яке поширюється з обох боків синапсу. У цьому процесі беруть участь потенціалзалежні іонні канали.
Ацетилхолін виділяється у незначній кількості навіть тоді, коли ПД нервовими волокнами не надходить. Навіть у такій кількості (кванти медіатора) він зумовлює незначну деполяризацію постсинаптичної мембрани (частки мВ), яка має назву мініатюрних потенціалів кінцевої платівки (МПКП).
Ацетилхолін, що виділився в синаптичну щілину, дуже швидко руйнується ферментом ацетилхолінестеразою. Внаслідок цього утворюється холін, який потрапляє у синаптичну бляшку та бере участь у створенні нових порцій ацетилхоліну.
Передача інформації в нервово-м'язовому синапсі відбувається в одному напрямку з певною затримкою (близько 0,5 мс). При занадто частих і тривалих подразненнях можуть наступати виснаження запасів ацетилхоліну та втома, є методи посилення або послаблення нерідачі імпульсів нервово-м'язових синапсів. Так, посилити передачу їх можна за допомогою замінників ацетилхоліну-холіноміметичних засобів (наприклад, карбахолін) або шляхом інактивації ацетилхолінестерази (езерин, фізостигмін). У цьому випадку медіатор не руйнується, його дія посилюється та продовжується.
Послабити або припинити передачу можна при введенні курареподібних речовин - міорелаксантів (наприклад, тубокурарину, диплацину). Ці препарати зв'язуються з холінорецепторами та блокують дію ацетилхоліну.
Фізіологія нервів.
Центральна нервова система людини складається з нервових клітин, кожна з яких має один аксон і багато дендритів. Нервові волокна поділяються на: м'якотні та безм'якотні. Всі вони мають шваннівську оболонку, а м'якотні, крім цього, вкриті ще мієліновою оболонкою, між якою є перехоплення Ранв'є, в яких мієлінова оболонка відсутня. Безм'якотні волокна мають малий діаметр, менше 1,3 мкм, невелику швидкість поширення імпульсу до 2 м/сек, тривалість ПД – 2 мсек. М'якотні виникли з безм'якотних, діаметр до 25 мкм, швидкість поширення збудження до 120 м/сек, тривалість ПД – 0,4-0,5 мсек.
Поширення ПД по безм'якотному волокну:німецький фізіолог Герман запропонував теорію «місцевих струмів», згідно з якою при поширенні збудження на мембрані нервового волокна між збудженою та незбудженою ділянкою виникає місцевий струм, який є подразником для незбудженої ділянки. Якщо його величина достатня виникнення ПД у сусідній ділянці, то ПД поширюється цей участок.
Розповсюдження місцевих струмів у безм'якотних нервових волокнах прямопропорційно опору мембрани і зворотнопропорційно опору внутрішньо- та позаклітинного середовища. Відстань на яку поширюються місцеві струми буде тим більшою, чим більша амплітуда ПД і чим менший пороговий потенціал.
У м'якотних волокнах збудження виникає при нанесенні роздратування в перехопленнях Ранв'є (теорія Тасакі) та поширюється по мембрані волокна сальтаторно (стрибкоподібно) (рис.1).
Малюнок 1. Проведення збудження по нервовому мієліновому волокну.
При цьому може охоплювати не тільки один, а й два перехоплення Ранв'є, що забезпечує надійність, а також збільшує швидкість поширення збудження та економічно вигідніша, оскільки на 1 імпульс у безм'якотному волокні енергія витрачається у 20 разів більше, ніж у м'якотному. Таким чином, швидкість і відстань, на яку поширюється збудження в безм'якотних волокнах пропорційна діаметру, опору мембрани та амплітуді ПД; у м'якотних прямопропорційна довжині міжперехоплювальних ділянок, які довжина тим більше, що більше діаметр волокна. Швидкість залежить від сили подразнення.
Теорію Германа було експериментально підтверджено.
Гельмгольц - визначив швидкість поширення імпульсу у жаби; Бабський – визначив швидкість поширення імпульсу в людини. Ерландер і Гассер класифікували різні нервові волокна на три групи А, В та С:
Типи нервових волокон
А - мієлінізовані, найбільшого діаметра, швидкість 120-70 м в сек, тривалість ПД - мінімальна, поділяються на підгрупи: альфа, бета, гама, дельта. Приклад – аксони мотонейронів.
В - мієлінізовані волокна, меншого діаметра, швидкість 3-18 м в сек, ПД більш тривалий. Приклад - прегангліонарні волокна симпатичної нервової системи.
С – немієлізовані нервові волокна, швидкість менше 2 м на сік, тривалість ПД максимальна. Приклад - постгангліонарні волокна парасимпатичної нервової системи.
Закони проведення порушення нервами.
I законанатомо-фізіологічний цілісність нервового волокна. Щоб збудження поширювалося нервовим волокном необхідно як його морфологічна цілісність, а й фізіологічна безперервність. Препарати для провідникової анестезії порушують фізіологічну безперервність тим, що інактивують натрієву проникність у нервових волокнах.
II законізольованого проведення збудження по нервовому волокну. У змішаному нерві збудження з одного нервового волокна не передається на сусідні, оскільки опір міжклітинної рідини менший за опір мембран сусідніх волокон. Цим забезпечується точність проведення інформації в нервових волокнах до структур, що іннервуються.
ІІІ закон двостороннє проведення збудження. Поширення ПД мембраною нервового волокна можливе в обидві сторони, оскільки будова мембрани протягом усього однаково. У той же час збудження не може повернутися в ділянку, де воно виникло, оскільки він перебуває у стані рефрактерності.
Парабіоз.Н.Є. Введенський, досліджуючи проходження імпульсу через відрізок нерва який впливаю хімічні чи наркотичні речовини (альтератори), спостерігав різке зниження лабільності. Парабіоз характеризується поступовим розвитком, у якому можна виділити чотири фази:
I Продромальна (не завжди проявляється, тому що дуже короткочасна) характеризується підвищенням збудливості, підвищенням лабільності.
II Зрівняльна – ефекти від сильних та слабких подразників урівноважуються.
III Парадоксальна – на сильні, або часті подразнення ефект буває меншим, ніж на слабкі чи рідкісні.
IV Гальмівна – ні сильні, ні слабкі роздратування викликають скорочення м'язів. Через уражену ділянку не проходять імпульси.
Якщо другий парою електродів вплинути на уражений ділянку, збудження буде, тобто. тканина ще жива.
Якщо зняти альтератор, то тканина повертається до початкового стану зворотному порядку фаз IV, III, II, I.
Парабіоз - це стійке збудження, що не поширюється.
Виникають менші потенціали по своїй амплітуді, а далі абортивні потенціали, не здатні поширюватися: зменшуються процеси Na-евої проникності, і збільшуються процеси Na-евой інактивації.
Нервово-м'язова передача.
Взаємодія людини із зовнішнім середовищем неможливо уявити без її м'язової системи. Вироблені рухи скелетною мускулатурою необхідні для виконання найпростіших переміщень тіла в просторі, складних маніпуляцій хірурга, стоматолога, вираження найтонших почуттів і думок за допомогою мови, міміки, жестів. Робота серця забезпечує кровопостачання всіх органів, робота гладких м'язів створює умови для нормального здійснення фізіологічних процесів, що забезпечують гомеостаз, практично у всіх системах: гастроінтестинальної, серцево-судинної, видільної, репродуктивної, дихальної. Провідна роль скелетної мускулатури також у виробництві тепла та підтримці температури тіла. М'язи - це «машини», що перетворюють хімічну енергію на механічну (роботу) та тепло. Маса м'язів більша за інші органи, 40-50% від маси тіла.
У природних умовах (у нашому організмі) збудження м'язового волокна (або декількох м'язових волокон, що становлять м'яз) виникає в результаті передачі збудження з нервового волокна на м'язову мембрану в місцях контакту нерва і м'язи: нервово-м'язових синапсах.
Механізм нервово-м'язової передачі
Синапси являють собою комунікаційні структури, які формуються закінченням нервового волокна та прилеглої до нього мембраною м'язового волокна (пресинаптичною нервовою та постсинаптичною м'язовою мембранами) (рис.2). 
Рис.2. Міоневральний синапс.
Коли нервовий імпульс досягає закінчення аксона, на деполяризованій пресинаптичній мембрані відкриваються потенціалзалежні Са2+ канали. Вхід Са2+ в аксональне розширення (пресинаптичну мембрану) сприяє вивільненню хімічних нейромедіаторів, що у вигляді везикул (бульбашок) із закінчення аксона. Медіатори (в нервово-м'язовому синапсі це завжди ацетилхолін) синтезуються в сомі нервової клітини і шляхом аксонального транспорту транспортуються до закінчення аксона, де виконують свою роль. Медіатор дифундує через синаптичну щілину та зв'язується зі специфічними рецепторами на постсинаптичній мембрані. Оскільки медіатором у нервово-м'язовому синапсі є ацетилхолін, то рецептори постсинаптичної мембрани називають холінорецепторами. Внаслідок цього процесу на постсинаптичній мембрані відкриваються хемочутливі Nа+-канали, виникає деполяризація, величина якої різна, і залежить від кількості виділеного медіатора. Найчастіше виникає локальний процес, який називають потенціалом кінцевої платівки (ПКП). При підвищенні частоти стимуляції нервового волокна, посилюється деполяризація пресинаптичної мембрани, а, отже, зростає кількість медіатора, що виділяється, і число активованих хемочутливих Nа+каналів на постсинаптичній мембрані. Таким чином, виникають ПКП, які по амплітуді деполяризації підсумовуються до порогового рівня, після чого на мембрані м'язового волокна, що оточує синапс, виникає ПД, який має здатність до поширення вздовж мембрани м'язового волокна. Чутливість постсинаптичної мембрани регулюється активністю ферменту – ацетилхолінестерази (АЦХ-Е), який гідролізує медіатор АЦХ на складові компоненти (ацетил та холін) та повертає назад – у пресинаптичну бляшку для ресинтезу. Без видалення медіатора на постсинаптичній мембрані розвивається тривала деполяризація, яка веде до порушення проведення збудження у синапсі – синаптичної депресії. Таким чином, синаптичний зв'язок забезпечує одностороннє проведення збудження з нерва на м'яз, проте на всі ці процеси витрачається час (синаптична затримка), що призводить до низької лабільності синапсу в порівнянні з нервовим волокном.
Таким чином, нервово-м'язовий синапсє «вигідним» місцем,куди можна вплинути фармакологічними препаратами, змінюючи чутливість рецептора, активність ферменту. Ці явища часто зустрічатимуться у практиці лікаря: наприклад, при отруєнні токсином ботулізму – блокується вивільнення медіатора АЦХ (розгладжування зморшок у косметичній медицині), блокада холіноререпторів (курареподібними препаратами, бунгаротоксином) порушує відкриття Nа+ каналів на постсинапти. Фосфоорганічні сполуки (безліч інсектицидів) порушує ефективність АЦХ-Е та викликає тривалу деполяризацію постсинаптичної мембрани. У клініці використовують специфічні блокатори нервово-м'язового проведення: блокада холінорецепторів курареподібними препаратами, сукцинілхоліном та іншими конкурентними інгібіторами, що витісняють АЦХ із холінорецептора. При захворюванні на міастенію через дефіцит холінорецепторів на постсинаптичній мембрані (через їх аутолітичне руйнування) виникає прогресуюча м'язова слабкість, аж до повної зупинки м'язових скорочень (зупинка дихання). У цьому випадку використовують блокатори АЦХ-Е, що призводить до збільшення тривалості зв'язування медіатора з меншою кількістю холінорецепторів та дещо збільшує амплітуду деполяризації постсинаптичної мембрани.
Фізіологія м'язів
Існує 3 типи м'язової тканини: поперечно-смугаста, серцева, гладка.
Скелетнам'язова тканина утворює великий обсяг соматичної мускулатури. Містить добре виражену впорядковану будову скоротливих білків як поперечної смугастість. Зв'язків між окремими м'язовими волокнами немає. Регуляція скорочень відбувається свідомо.
Серцевам'язмістить також поперечну смугастість, але є функціональним синцитієм. Завдяки наявності пейсмекерних клітин, має здатність генерувати спонтанні нервові імпульси, що забезпечують серцеві скорочення.
У гладкихм'язах немає поперечної смугастість, яка б надавала певні фізичні та фізіологічні властивості цієї тканини. Так, на відміну від скелетної, якій властива висока еластичність, у гладкому м'язі більше виражена властивість пластичності, що зумовлено відсутністю чіткої впорядкованості міофіламентів актину та міозину. На відміну від регулярної саркомірної структури скелетних та серцевих м'язів, тонкі філаменти гладких м'язів приєднані до структур у цитоплазмі, званих щільними тільцями(Прикріплювальними бляшками сарколеми), що складаються з білка десміну. Гладкі м'язи бувають вісцеральні (мультіунітарні) та поліелементні (унітарні).
Вісцеральнімістять містки-щілинні контакти з низьким опором електричного струму - синцитій. Вісцеральні м'язи зустрічаються в стінках порожнистих органів (кишковик, матка, сечоводи, сечовий міхур).
Поліелементнігладкі м'язи складаються з окремих м'язових одиниць без сполучних містків, тому для них характерні точні, чітко дозовані скорочення, подібно до скелетних. Однак, скорочення цих м'язів не можна контролювати, на відміну від скелетних (м'язи райдужної оболонки ока, циліарного тіла, насіннєвих проток, артерії). Завдяки своїй будові (вісцеральні)мають велику пластичність, проте при певному граничному ступені розтягування здатні деполяризуватися і скорочуватися (саморегуляція). Наприклад, тиск усередині стінок сечового міхура мало змінюється при відносно невеликому його розтягуванні, якщо розтягування зростає різко – відбувається скорочення м'язів детрузора – евакуація сечі навіть у випадках, коли іннервація порушена. Поліелементнігладкі м'язи мають більш потужну (щільну) іннервацію та слабкий розвиток міжклітинних контактів (нексусів). Тонус цих м'язів та його коливання мають нейрогенну природу. Мають парасимпатичні та симпатичні, а також метасимпатичні волокна. Будова нервових закінчень у гладкому м'язі відрізняється від будови нервово-м'язового синапсу в скелетному м'язі. У гладкому м'язі немає кінцевих платівок та окремих нервових закінчень. По всій довжині розгалужень адренергічних та холінергічних нервових волокон є потовщення, які називаються варикозами. Вони містять гранули з медіатором, що виділяється з кожної варикози. Клітини, позбавлені безпосередніх контактів з варикозами, активуються ПД, що розповсюджуються через нексуси сусідні клітини. По ходу проходження нервового волокна м'язові клітини можуть збуджуватися або гальмуватися (стимуляція адренергічних волокон зменшує, а холінергічних – збільшує м'язову активність, в інших, наприклад у судинах, норадреналін – посилює, а ацетилхолін – зменшує м'язовий тонус. Іонна природа гладко-м'язової клітини Основну роль у механізмі генерації ПД грають іони Са2+, але цими каналами можуть всередину клітини рухатися й інші двовалетні іони Ва2+, Mg2+ Вхід Са2+ в клітину необхідний для підтримки тонусу м'язів і розвитку скорочення, тому блокування Са2+ каналів гладких м'язів призводить до обмеження надходження цього іону до цитоплазми міоцитів внутрішніх органів та судин, що широко використовується в практичній медицині для корекції моторної функції ШКТ та тонусу судин.
Регулювання м'язових скорочень залежить від обсягу іннервації. Аксон спинно-мозкового (рухового) мотонейрона розгалужується на кілька терміналей (гілочок), кожна з яких підходить до одного м'язового волокна. Тому в цілісному організмі при збудженні 1 нервового волокна скорочується група м'язів. Сукупність рухового нейрона та м'язових волокон, які він іннервує називають руховою одиницею. Кількість м'язових волокон, що входять у рухову одиницю, по-різному і залежить від функції, яку реалізує кожен конкретний м'яз в організмі. У м'язах очей, кисті руки, які забезпечують високоточні, координовані рухи, 1 рухова одиниця містить 3-5 м'язових волокон. Двигуни м'язів спини, стегна - складаються з декількох сотень м'язових волокон, які регулюються одним мотонейроном.
Повернемося до мембрани м'язового волокна, де поширюється ПД, що виник у результаті «успішного» проведення порушення через синапс. Продовженням мембрани м'язового волокна є саркотубулярна система, що утворює поперечні інвагінації (вп'ячування) (Т-система поперечних трубочок). Система Т-трубочок виконує кілька важливих функцій: є внутрішньоклітинним депо іонів Са2+; повідомляється із позаклітинною рідиною і таким чином регулюється вміст Са2+ у ній; містить потенційно чутливі Nа+ канали, які дають можливість ПД поширюватися як вздовж, так і вглиб м'язового волокна. Система Т-трубочок забезпечує швидке та узгоджене збудження м'язової клітини, оскільки поширення деполяризації по Т-системі пов'язане з виходом Са2+ з саркоплазматичних цистерн ретикулума, забезпечує ініціацію скорочення м'яза. Через Т-трубочки може відбуватися виділення продуктів обміну (наприклад, молочної кислоти) з м'язової клітини інтерстицій (міжклітинний простір) і далі в кров. Великий обсяг клітини скелетного м'яза неможливо було б активувати швидко, якби Са2+ надходив із позаклітинного середовища. Нагромадженню Са2+ у саркоплазматичному ретикулумі сприяє білок (кальсеквестрин), який неміцно зв'язує Са2+ у ретикулумі. Вивільнення Са2+ із саркоплазматичного ретикулуму в скелетних м'язах відбувається завдяки взаємодії Т трубочок та кінцевих цистерн (тріада).
Іони Са2+ потрапивши до саркоплазми ініціюють скорочення, зв'язуючись з білком тропоніном – блокатором актину у спокої (рис.3).

Малюнок. 3. Механізм м'язового скорочення.
Тонка нитка актину складається з 2 тяжів фібрилярного актину, на якому нанизані окремі глобули мономеру актину (як намисто). Актин містить активні центри (сайти) зв'язування з міозином, які у стані спокою заблоковані тропоніном. Тропонії пригнічує АТФ-азну активність міозину, що унеможливлює розщеплення АТФ і м'язові волокна перебувають у розслабленому стані. Великий скорочувальний білок - міозин, що складається з шести поліпептидних ланцюгів, покладених попарно. 2 з них - важкі ланцюги міоглобіну, що мають властивість ферментів. Пов'язаний із трононіном Са2+ звільняє активні центри актину для контакту з міозином. У присутності актину міозиновий фермент (глобулярна головка) руйнує АТФ і взаємодіє з тонкими нитками актину, створюючи рушійну силу скорочення – утворюючи поперечні містки («гребки») і м'яз скорочується (скорочується). Таким чином, АТФ забезпечує скорочення енергією, гідролізуючись на глобулярних головках міозину. Енергія (Е), яка вивільняється при гідролізі АТФ, перетворюється в силу скорочення за рахунок конформаційних (просторових) змін у міозині (робочий хід – утворення поперечних гребкових містків) – це тепло активації, що виділяє при зв'язуванні актину та міозину. АДФ, пов'язана з міозином, зменшує спорідненість поперечного містка до активного центру актину, що ініціює наступну фазу – розслаблення м'язів. За рахунок гідролізу АТФ виділяється Е (тепло укорочення), яка йде на:
1) роботу Са-АТФ-ази (активація насоса відбувається за рахунок неорганічного фосфату, що утворюється при гідролізі АТФ),
2) за рахунок Е відбувається відкачування проти градієнта концентрації Са2+ у саркоплазматичний ретикулум (активний транспорт).
3) АТФ у м'язовій клітині забезпечує роботу Na+-К-АТФ-ази, що забезпечує видалення Na+ з клітини та відновлення потенціалу спокою (а, отже, і збудливості) м'язової клітини.
АТФ забезпечує процеси, як скорочення, і розслаблення. Якщо Са2+ не транспортуватиметься назад у СПР, то розслаблення не настане, розвивається ригідність м'яза (при трупному задубінні), або в живому організмі – при посттетанічній контрактурі – накопичення Са2+ у саркоплазмі ініціює тривале м'язове скорочення поза надходженням ПД через синапс на мембрану м'язового.
Гладкий м'яз містить також тропоміозин, але не має тропоніну, співвідношення актину до міозину 14-16 до 1, порівняйте у скелетних співвідношення актину до міозину 2 до 1. Гладкий м'яз має щілинні контакти – це містки, що з'єднують мембрани сусідніх клітин. Регуляція скорочувальної активності гладких м'язів відбувається завдяки зв'язуванню Са2+ з кальмодуліном, що активує кіназу легкого ланцюга міозину, що призводить до гідролізу АТФ та запускає цикл утворення поперечних містків.
ПД скелетного м'яза триває близько 2-4 мс і проходить по мембрані м'язового волокна зі швидкістю близько 5 м/с. 1 ПД викликає одиночне м'язове скорочення, яке починається через 2 мс після початку деполяризації мембрани (латентний період) та завершується скорочення майже одночасно з реполяризацією. Тривалість одиночного скорочення різна і від типу м'язової тканини. При частих стимулах розвивається сумарне м'язове скорочення всіх м'язових волокон, що мають різний опір мембран до електричного струму. Однак незначні відмінності порогів збудження забезпечують синхронність м'язового скорочення цілого м'яза. Наявність абсолютного рефрактерного періоду близько 1-3 мс, що зумовлює виникнення різних видів тетанусів (сумарних м'язових скорочень). Існує зубчастий та гладкий тетануси. Частота стимуляції м'яза у розвиток гладкого тетануса має бути вищою, ніж у розвитку зубчастого. Стимул повинен потрапляти у фазу укорочення м'яза, якщо ж м'язи почала розслаблятися, а ми його стимулюємо, то отримуємо – зубчастий тетанус. Мінімальний проміжок часу між послідовними ефективними стимулами під час тетанусу не може бути меншим від рефрактерного періоду, який приблизно відповідає тривалості ПД. Оскільки м'язи складаються з м'язових волокон з різним рівнем збудливості, є певна залежність між величиною стимулу і реакцією у відповідь. Збільшення сили скорочення можливе до певної межі, після якого амплітуда скорочення залишається незмінною зі збільшенням амплітуди стимулу (треба зазначити, що в м'язі підсумовуються не ПД, а скорочення). При цьому всі волокна, що входять до складу м'яза, беруть участь у скороченні.
В організмі людини є швидкі, фазні м'язові волокна (білі), тривалість скорочення яких до 7,5 мс, і повільні, тонічні (червоні), які забезпечують сильні та потужні рухи, що тривають до 100 мс. Червоні (тонічні) мають багато волокон міозину типу I, що відрізняються низькою активністю АТФ-ази міозину. Швидкість розщеплення АТФ є фактором, що визначає частоту гребкових рухів, і таким чином, швидкість ковзання ниток актину вздовж міозину. З СПР Са2+ транспортується повільно, висока окислювальна здатність, багато капілярів, багато міоглобіну у структурі міозину (важкі ланцюги), багато мітохондрій. На стимул реагують повільно, мають тривалий латентний період скорочення, тому здатні до тривалих, повільних, тонічних скорочень, більш резистентні до втоми. Головна функція – підтримання становища тіла. Білі м'язи містять волокна міозину ІІ типу. Гліколітичний тип окислення, мало міоглобіну, мітохондрій, це волокна великого діаметру з високою активністю АТФ-ази міозину, здатні розвинути значну силу, але швидко втомлюються.
Сила м'язіввизначається тим максимальним вантажем, який м'яз може трохи підняти. Сила різних м'язів неоднакова. Для порівняння сили різних м'язів максимальний вантаж, який м'яз може підняти ділять на число квадратних сантиметрів її фізіологічного поперечного перерізу. Силові характеристики вищі у м'язів з перистим (косим) розташуванням волокон, при цьому фізіологічний переріз більший за геометричний поперечний переріз. Сума поперечного перерізу який завжди збігається з фізіологічним перерізом м'яза (тільки при паралельному розташуванні волокон.
Сила скороченняізольованого скелетного м'яза за інших рівних умов залежить від вихідної довжини м'яза. Помірне розтягнення м'яза призводить до того, що сила, що розвивається, зростає в порівнянні з силою, яку розвиває нерозтягнутий м'яз. Відбувається підсумовування пасивної напруги, обумовленої наявністю еластичних компонентів м'яза, та активного скорочення (перехід до правила середніх навантажень – фізіологічний механізм цього закону). Здатність здійснювати роботувизначається добутком величини піднятого вантажу на висоту підйому. Величина роботи м'яза поступово збільшується зі збільшенням маси вантажу, що піднімається, але до певної межі, після якого збільшення маси вантажу призводить до зниження величини роботи, так як висота підйому вантажу різко падає. Отже, максимальна робота здійснюється м'язом при середніх величин навантажень.Сила скорочення та робота не залишаються постійними при статичній та динамічній роботі. Через війну тривалої діяльності працездатність скелетної мускулатури знижується. Це явище називають втомою.При цьому знижується сила скорочень, збільшується латентний період скорочення та період розслаблення. Статичний режим роботи більш стомливий, ніж динамічний (чому пояснити). Накопичуються продукти процесів окислення - молочна піровіноградна кислота, які знижують можливість генерування ПД. Порушуються процеси ресинтезу АТФ та креатинфосфату, необхідні для енергетичного забезпечення м'язового скорочення. У природних умовах м'язова втома при статичній роботі переважно визначається неадекватним регіонарним кровотоком. Виникає «кисневе голодування» та втома прогресивно наростає.
У 1903 році Сєченов – відновлення працездатності стомленого м'яза значно прискорюється при виконанні роботи іншим м'язом у період відпочинку стомленого – такий відпочинок називають активним.Проте високий рівень розумової діяльності прискорює процеси розвитку втоми у м'язовій системі (втома нервових центрів).