Основна функція мембрани клітки. Клітинна мембрана.
Усі живі організми Землі складаються з клітин, кожна клітина оточена захисної оболонкою – мембраною. Однак функції мембрани не обмежуються захистом органоїдів та відділенням однієї клітини від іншої. Клітинна мембрана є складним механізмом, що безпосередньо бере участь у розмноженні, регенерації, харчуванні, диханні та багатьох інших важливих функціях клітини.
Термін "клітинна мембрана" використовується вже близько ста років. Саме слово "мембрана" у перекладі з латині означає "плівка". Але у випадку клітинної мембраною правильніше буде говорити і сукупності двох плівок, з'єднаних між собою певним чином, причому, різні сторони цих плівок мають різні властивості.
Клітинна мембрана (цитолема, плазмалема) – це тришарова ліпопротеїнова (жиро-білкова) оболонка, що відокремлює кожну клітину від сусідніх клітин та навколишнього середовища, та здійснює керований обмін між клітинами та навколишнім середовищем.
Вирішальне значення в цьому визначенні має не те, що клітинна оболонка відокремлює одну клітину від іншої, а те, що вона забезпечує її взаємодію іншими клітинами та навколишнім середовищем. Мембрана – дуже активна, постійно діюча структура клітини, яку природою покладено безліч функцій. З нашої статті ви дізнаєтеся все про склад, будову, властивості та функції клітинної мембрани, а також про ту небезпеку, яку становлять для здоров'я людини порушення в роботі клітинних мембран.
Історія дослідження клітинної мембрани
1925 року двоє німецьких учених, Гортер і Грендель, змогли провести найскладніший експеримент над червоними кров'яними тільцями людської крові, еритроцитами. За допомогою осмотичного удару дослідники отримали так звані тіні - порожні оболонки еритроцитів, потім склали їх в одну стопку і виміряли площу поверхні. Наступним кроком стало обчислення кількості ліпідів у клітинній мембрані. За допомогою ацетону вчені виділили ліпіди з «тіней» і визначили, що їх вистачає на подвійний суцільний шар.
Однак у ході експерименту було допущено дві грубі помилки:
Використання ацетону не дозволяє виділити з мембрани абсолютно всі ліпіди;
Площа поверхні "тіней" була вирахована за сухою вагою, що теж неправильно.
Оскільки перша помилка давала мінус у розрахунках, а друга – плюс, загальний результат виявився напрочуд точним, і німецькі вчені принесли у науковий світ найважливіше відкриття – ліпідний бислой клітинної мембрани.
У 1935 році інша пара дослідників, Даніеллі та Доусон, після довгих експериментів над біліпідними плівками дійшли висновку про присутність у клітинних мембранах білків. Інакше ніяк не можна було пояснити, чому ці плівки мають такий високий показник поверхневого натягу. Вчені представили до уваги громадськості схематичну модель клітинної мембрани, схожу на сендвіч, де роль шматочків хліба грають однорідні ліпідно-білкові шари, а між ними замість олії – порожнеча.
У 1950 році за допомогою першого електронного мікроскопа теорію Даніеллі-Доусона вдалося частково підтвердити - на мікрофотографіях клітинної мембрани були чітко видно два шари, що складаються з ліпідних та білкових головок, а між ними прозорий простір, заповнений лише хвостиками ліпідів та білків.
У 1960 році, керуючись цими даними, американський мікробіолог Дж. Робертсон розробив теорію про тришарову будову клітинних мембран, яка довгий час вважалася єдино правильною. Проте з розвитком науки народжувалося дедалі більше сумнівів щодо однорідності цих верств. З точки зору термодинаміки така будова вкрай невигідна – клітинам було б дуже складно транспортувати речовини всередину та назовні через весь бутерброд. Крім того, було доведено, що клітинні мембрани різних тканин мають різну товщину та спосіб кріплення, що зумовлено різними функціями органів.
1972 року мікробіологи С.Д. Сінгер та Г.Л. Ніколсон змогли пояснити всі нестикування теорії Робертсона за допомогою нової, рідинно-мозаїчної моделі клітинної мембрани. Вчені встановили, що мембрана неоднорідна, асиметрична, наповнена рідиною, та її клітини перебувають у постійному русі. А білки, що входять до її складу, мають різну будову та призначення, крім того, вони по-різному розташовуються щодо біліпідного шару мембрани.
У складі клітинних мембран присутні білки трьох видів:
Периферичні – кріпляться поверхні плівки;
Напівінтегральні– частково проникають усередину біліпідного шару;
Інтегральні – повністю пронизують мембрану.
Периферичні білки пов'язані з головками мембранних ліпідів за допомогою електростатичної взаємодії, і вони ніколи не утворюють суцільний шар, як прийнято було вважати раніше. для кількох важливих функцій, про які ви дізнаєтесь далі.

Клітинна мембрана виконує такі функції:
Бар'єрну – проникність мембрани для різних типів молекул неоднакова. Щоб уникнути оболонку клітини, молекула повинна мати певний розмір, хімічні властивості та електричний заряд. Шкідливі або невідповідні молекули завдяки бар'єрній функції клітинної мембрани просто не можуть проникнути всередину клітини. Наприклад, за допомогою реакції пероксису мембрана захищає цитоплазму від небезпечних для неї пероксидів;
Транспортну – крізь мембрану проходить пасивний, активний, регульований та вибірковий обмін. Пасивний обмін підходить для жиророзчинних речовин та газів, що складаються з дуже маленьких молекул. Такі речовини проникають усередину і виходять із клітини без витрат енергії, вільно методом дифузії. Активна транспортна функція клітинної мембрани задіюється тоді, коли в клітину або з неї потрібно провести необхідні речовини, що важко транспортуються. Наприклад, які мають великий розмір молекул, або нездатні перетнути біліпідний шар через гідрофобність. Тоді починають працювати білки-насоси, у тому числі АТФаза, яка відповідає за всмоктування в клітину іонів калію та викидання з неї іонів натрію. Регульований транспортний обмін необхідний здійснення функцій секреції і ферментації, наприклад, коли клітини виробляють і виділяють гормони чи шлунковий сік. Всі ці речовини виходять із клітин через спеціальні канали та в заданому обсязі. А виборча транспортна функція пов'язана з тими самими інтегральними білками, які пронизують мембрану і є каналом для входу і виходу строго певних типів молекул;
Матричну – клітинна мембрана визначає та фіксує розташування органоїдів щодо один одного (ядра, мітохондрій, хлоропластів) та регулює взаємодію між ними;
Механічну забезпечує обмеження однієї клітини від іншої, і, в той же час, - правильне з'єднання клітин в однорідну тканину і стійкість органів до деформації;
Захисну – як у рослин, так і у тварин, клітинна мембрана є основою для побудови захисного каркасу. Прикладом можуть бути тверда деревина, щільна шкірка, колючі шипи. У тваринному світі також багато прикладів захисної функції клітинних мембран - черепаший панцир, хітинова оболонка, копита та роги;
Енергетичну – процеси фотосинтезу та клітинного дихання були б неможливі без участі білків клітинної мембрани, адже саме за допомогою білкових каналів клітини обмінюються енергією;
Рецепторну - білки, вбудовані в клітинну мембрану, можуть мати ще одну важливу функцію. Вони служать рецепторами, завдяки яким клітина отримує сигнал від гормонів та нейромедіаторів. А це, у свою чергу, необхідно для проведення нервових імпульсів та нормальної течії гормональних процесів;
Ферментативну – ще одна важлива функція, властива деяким білкам клітинних мембран. Наприклад, в епітелії кишківника за допомогою таких білків синтезуються травні ферменти;
Біопотенційну- Концентрація іонів калію всередині клітини значно вище, ніж зовні, а концентрація іонів натрію, навпаки, зовні більше, ніж усередині. Цим і пояснюється різниця потенціалів: усередині клітини заряд негативний, зовні позитивний, що сприяє руху речовин усередину клітини та назовні при будь-якому з трьох типів обміну – фагоцитозі, піноцитозі та екзоцитозі;
Маркувальну - на поверхні клітинних мембран є так звані «ярлики» - антигени, що складаються з глікопротеїнів (білків з приєднаними до них розгалуженими олігосахаридними бічними ланцюгами). Оскільки бічні ланцюги можуть мати безліч конфігурацій, кожен тип клітин отримує свій унікальний ярлик, який дозволяє іншим клітинам організму впізнавати їх «в обличчя» і правильно на них реагувати. Ось чому, наприклад, імунні клітини людини, макрофаги, легко розпізнають чужинця, що проник в організм (інфекцію, вірус) і намагаються його знищити. Те саме відбувається з хворими, що мутували і старими клітинами – ярлик на їхній клітинній мембрані змінюється, і організм позбавляється їх.
Клітинний обмін відбувається через мембрани і може здійснюватися за допомогою трьох основних типів реакцій:
Фагоцитоз - клітинний процес, при якому вбудовані в мембрану клітини-фагоцити захоплюють та перетравлюють тверді частинки поживних речовин. У людському організмі фагоцитоз здійснюється мембранами двох типів клітин: гранулоцитів (зернистих лейкоцитів) та макрофагів (імунних клітин-убивць);
Піноцитоз - процес захоплення поверхнею клітинної мембрани стикаються з нею молекул рідини. Для харчування за типом піноцитозу клітина вирощує на своїй мембрані тонкі пухнасті вирости у формі вусиків, які ніби оточують крапельку рідини, і виходить пляшечку. Спочатку цей пляшечку випинається над поверхнею мембрани, а потім «ковтається» - ховається всередину клітини, і його стінки зливаються вже з внутрішньою поверхнею клітинної мембрани. Піноцитоз проходить майже у всіх живих клітинах;
Екзоцитоз - зворотний процес, при якому всередині клітини утворюються бульбашки з секреторною функціональною рідиною (ферментом, гормоном), і її необхідно вивести з клітини в навколишнє середовище. Для цього бульбашка спочатку зливається з внутрішньою поверхнею клітинної мембрани, потім випинається назовні, лопається, вириває вміст і знову зливається з поверхнею мембрани, цього разу вже із зовнішнього боку. Екзоцитоз проходить, наприклад, у клітинах кишкового епітелію та кори надниркових залоз.
Знайшли помилку у тексті? Виділіть її та ще кілька слів, натисніть Ctrl+Enter

Клітинні мембрани містять ліпіди трьох класів:
Фосфоліпіди;
Гліколіпіди;
Холестерол.
Фосфоліпіди (комбінація жирів і фосфору) і гліколіпіди (комбінація жирів і вуглеводів), у свою чергу, складаються з гідрофільної головки, від якої відходять два довгі гідрофобні хвостики. А ось холестерол іноді займає простір між цими двома хвостиками і не дає їм згинатися, що робить мембрани деяких клітин жорсткими. Крім того, молекули холестеролу впорядковують структуру клітинних мембран і перешкоджають переходу полярних молекул з однієї клітини до іншої.
Але найважливішою складовою, як видно з попереднього розділу про функції клітинних мембран, є білки. Їх склад, призначення та розташування дуже різноманітні, але є щось спільне, що їх об'єднує: навколо білків клітинних мембран завжди розташовані аннулярні ліпіди. Це особливі жири, які чітко структуровані, стійкі, мають у своєму складі більше насичених жирних кислот і виділяються з мембран разом з «підшефними» білками. Це свого роду персональна захисна оболонка для білків, без якої вони просто не працювали б.
Структура клітинної мембрани тришарова. Посередині пролягає відносно однорідний рідкий шар біліпіду, а білки покривають його з обох боків подобою мозаїки, частково проникаючи в товщу. Тобто неправильно було б думати, що зовнішні білкові шари клітинних мембран безперервні. Білки крім своїх складних функцій потрібні в мембрані для того, щоб пропускати всередину клітин і транспортувати з них назовні ті речовини, які не здатні проникнути крізь жировий шар. Наприклад, іони калію та натрію. Їх передбачені спеціальні білкові структури – іонні канали, докладніше про які ми розповімо далі.
Якщо поглянути на клітинну мембрану через мікроскоп, то можна побачити шар ліпідів, утворений дрібними кулястими молекулами, яким, як по морю, плавають великі білкові клітини різної форми. Такі самі мембрани ділять внутрішній простір кожної клітини на відсіки, у яких затишно розташовуються ядро, хлоропласти і мітохондрії. Якби не було всередині клітини окремих «кімнат», органоїди злиплися б один з одним і не змогли б виконувати свої функції правильно.
Клітина – це структурована та відмежована за допомогою мембран сукупність органоїдів, яка бере участь у комплексі енергетичних, метаболічних, інформаційних та репродуктивних процесів, що забезпечують життєдіяльність організму.
Як видно з цього визначення, мембрана є найважливішою функціональною складовою будь-якої клітини. Її значення так само велике, як значення ядра, мітохондрій та інших клітинних органел. А унікальні властивості мембрани обумовлені її будовою: вона складається з двох плівочок, зліплених один з одним особливим чином. Молекули фосфоліпідів у мембрані розташовані гідрофільними головками назовні, а гідрофобними хвостами усередину. Тому один бік плівочки змочується водою, а інший – ні. Так от, ці плівочки з'єднуються один з одним незмочуваними сторонами всередину, утворюючи біліпідний шар, оточений молекулами білків. Це і є та сама «бутербродна» будова клітинної мембрани.
Іонні канали клітинних мембран
Розглянемо докладніше принцип роботи іонних каналів. Навіщо вони потрібні? Справа в тому, що крізь ліпідну мембрану безперешкодно можуть проникати лише жиророзчинні речовини – це гази, спирти та самі жири. Так, наприклад, у червоних кров'яних тільцях постійно відбувається обмін кисню та вуглекислого газу, і для цього нашому організму не доводиться вдаватися до жодних додаткових хитрощів. Але як бути, коли виникає необхідність у транспортуванні крізь клітинну мембрану водних розчинів, таких, як солі натрію і калію?
Прокласти в біліпідному шарі шлях для таких речовин було б неможливо, оскільки отвори тут же затягнулися б і злиплися назад, така вже структура будь-якої жирової тканини. Але природа, як завжди, знайшла вихід із ситуації, і створила спеціальні білкові транспортні структури.
Існує два типи провідних білків:
Транспортери – напівінтегральні білки-насоси;
Каналоформери – інтегральні білки.
Білки першого типу частково занурені в біліпідний шар клітинної мембрани, а головкою виглядають назовні, і в присутності потрібної речовини вони починають поводитися як насос: притягують молекулу і всмоктують її всередину клітини. А білки другого типу, інтегральні, мають витягнуту форму і розташовуються перпендикулярно до біліпідного шару клітинної мембрани, пронизуючи її наскрізь. По них, як по тунелях, у клітину і з клітини рухаються речовини, нездатні проходити крізь жир. Саме через іонні канали всередину клітини проникають іони калію та накопичуються у ній, а іони натрію, навпаки, виводяться назовні. Виникає різниця електричних потенціалів, така необхідна для правильної роботи всіх клітин нашого організму.
Найважливіші висновки про будову та функції клітинних мембран

Теорія завжди виглядає цікавою та перспективною, якщо її можна з користю застосувати на практиці. Відкриття будови та функцій клітинних мембран людського організму дозволило вченим зробити справжній прорив у науці в цілому, і в медицині зокрема. Ми не випадково так докладно зупинилися на іонних каналах, адже саме тут криється відповідь на одне з найважливіших питань сучасності: чому люди все частіше хворіють на онкологію?
Рак щорічно забирає близько 17 мільйонів життів у всьому світі, і є четвертою за частотою причиною всіх смертей. За даними ВООЗ, захворюваність на онкологію неухильно збільшується, і до кінця 2020 року може досягти 25 мільйонів на рік.
Чим пояснюється реальна епідемія раку, і тут функції клітинних мембран? Ви скажете: причина у поганій екологічній обстановці, неправильному харчуванні, шкідливих звичках та важкій спадковості. І, звичайно, матимете рацію, але якщо говорити про проблему більш предметно, то причина в закисленості людського організму. Наведені вище негативні фактори призводять до порушення роботи клітинних мембран, пригнічують дихання та харчування.
Там, де має бути плюс, утворюється мінус і клітина не може нормально функціонувати. А ось раковим клітинам не потрібні ні кисень, ні лужне середовище – вони здатні використовувати анаеробний тип харчування. Тому в умовах кисневого голодування і рівня pH, що зашкалює, здорові клітини мутують, бажаючи пристосуватися до навколишнього середовища, і стають раковими клітинами. Так людина і хворіє на онкологію. Щоб цього уникнути, потрібно лише вживати достатню кількість чистої води щодня, і відмовитися від канцерогенів у їжі. Але, як правило, люди чудово знають про шкідливі продукти та потребу в якісній воді, і нічого не роблять – сподіваються, що біда обійде їхньою стороною.
Знаючи особливості будови та функцій клітинних мембран різних клітин, лікарі можуть використовувати ці відомості для надання спрямованого, адресного терапевтичного впливу на організм. Багато сучасних лікарських препаратів, потрапляючи до нашого тіла, шукають необхідну «мішень», якою можуть бути іонні канали, ферменти, рецептори і біомаркери клітинних мембран. Такий спосіб лікування дозволяє досягти більш високих результатів за мінімальних побічних ефектів.
Антибіотики останнього покоління при попаданні в кров не вбивають усі клітини поспіль, а шукають саме клітини збудника, орієнтуючись на маркери у його клітинних оболонках. Нові препарати проти мігрені, триптани, звужують лише запалені судини головного мозку, при цьому майже не впливаючи на серце і периферичну кровоносну систему. І дізнаються вони про потрібні судини саме по білках їх клітинних мембран. Таких прикладів безліч, тому можна з упевненістю сказати, що знання про будову та функції клітинних оболонок лежить в основі розвитку сучасної медичної науки, і рятує мільйони життів щороку.
Автор статті: доктор медичних наук, лікар-терапевт Мочалов Павло Олександрович
Клітина- це як рідина, ферменти та інші речовини, а й високоорганізовані структури, звані внутрішньоклітинними органелами. Органели для клітини не менш важливі за її хімічні складові. Так, за відсутності таких органел, як мітохондрії, запас енергії, витягнутої з поживних речовин, одразу зменшиться на 95%.
Більшість органел у клітці покриті мембранами, що складаються в основному з ліпідів та білків. Розрізняють мембрани клітин, ендоплазматичного ретикулуму, мітохондрій, лізосом, апарату Гольджі.
Ліпідинерозчинні у воді, тому в клітині вони створюють бар'єр, що перешкоджає руху води та водорозчинних речовин з одного компартменту до іншого. Молекули білка, проте, роблять мембрану проникною різних речовин з допомогою спеціалізованих структур, званих порами. Багато інших мембранних білків є ферментами, що каталізують численні хімічні реакції, які будуть розглянуті в наступних розділах.
Клітинна (або плазматична) мембранає тонкою, гнучкою і еластичною структурою товщиною всього 7,5-10 нм. Вона складається в основному з білків та ліпідів. Приблизне співвідношення її компонентів таке: білки – 55%, фосфоліпіди – 25%, холестерол – 13%, інші ліпіди – 4%, вуглеводи – 3%.
Ліпідний шар клітинної мембраниперешкоджає проникненню води. Основу мембрани становить ліпідний бішар - тонка ліпідна плівка, що складається з двох моношарів і повністю покриває клітину. По всій мембрані розташовуються білки як великих глобул.
Ліпідний бислойскладається головним чином молекул фосфоліпідів. Один кінець такої молекули є гідрофільним, тобто. розчинним у воді (у ньому розташована фосфатна група), інший - гидрофобным, тобто. розчинним лише у жирах (на ньому знаходиться жирна кислота).
Завдяки тому, що гідрофобна частина молекули фосфоліпідувідштовхує воду, але притягується до подібних частин таких молекул, фосфоліпіди мають природну властивість прикріплюватися один до одного в товщі мембрани, як показано на рис. 2-3. Гідрофільна частина з фосфатною групою утворює дві мембранні поверхні: зовнішню, яка контактує із позаклітинною рідиною, та внутрішню, яка контактує із внутрішньоклітинною рідиною.
Середина ліпідного шарунепроникна для іонів та водних розчинів глюкози та сечовини. Жиророзчинні речовини, включаючи кисень, вуглекислий газ, алкоголь, навпаки, легко проникають через область мембрани.
Молекулихолестеролу, що входить до складу мембрани, за природою також відносяться до ліпідів, оскільки їх стероїдна угруповання має високу розчинність у жирах. Ці молекули розчинені в ліпідному бішарі. Їхнє головне призначення - регуляція проникності (або непроникності) мембран для водорозчинних компонентів рідких середовищ організму. Крім того, холестерол – основний регулятор в'язкості мембрани.
Білки клітинних мембран. На малюнку в ліпідному бісла видно глобулярні частинки - це мембранні білки, більшість яких є глікопротеїнами. Розрізняють два типи мембранних білків: (1) інтегральні, які пронизують мембрану наскрізь; (2) периферичні, які виступають лише над однією її поверхнею, не досягаючи іншою.
Багато інтегральних білківформують канали (або пори), через які у внутрішньо-і позаклітинну рідину можуть дифундувати вода та водорозчинні речовини, особливо іони. Завдяки вибірковості дії каналів одні речовини дифундують краще за інші.
Інші інтегральні білкифункціонують як білки-переносники, здійснюючи транспорт речовин, для яких ліпідний бислой непроникний. Іноді білки-переносники діють у протилежному напрямку дифузії, такий транспорт називають активним. Деякі інтегральні білки є ферментами.
Інтегральні білки мембраниможуть служити рецепторами для водорозчинних речовин, включаючи пептидні гормони, оскільки мембрана для них непроникна. Взаємодія білка-рецептора з певним лігандом призводить до конформаційних змін молекули білка, що, своєю чергою, стимулює ферментативну активність внутрішньоклітинного сегмента білкової молекули або передачу сигналу від рецептора всередину клітини за допомогою вторинного посередника. Таким чином, інтегральні білки, вбудовані в клітинну мембрану, залучають її в процес передачі інформації про довкілля всередину клітини.
Молекули периферичних мембранних білківчасто бувають пов'язані з інтегральними білками. Більшість периферичних білків є ферментами чи відіграють роль диспетчера транспорту речовин через мембранні пори.
_Теми _
4.1. Загальна характеристика мембран. Будова та склад мембран
4.2. Транспорт речовин через мембрани
4.3. Трансмембранна передача сигналів _
Цілі вивчення Вміти:
1. Інтерпретувати роль мембран у регуляції метаболізму, транспортуванні речовин у клітину та видаленні метаболітів.
2. Пояснювати молекулярні механізми дії гормонів та інших сигнальних молекул на органи-мішені.
Знати:
1. Будова біологічних мембран та його роль обміні речовин та енергії.
2. Основні методи перенесення речовин через мембрани.
3. Головні компоненти та етапи трансмембранної передачі сигналів гормонів, медіаторів, цитокінів, ейкозаноїдів.
ТЕМА 4.1. ЗАГАЛЬНА ХАРАКТЕРИСТИКА МЕМБРАН.
БУДОВА ТА СКЛАД МЕМБРАН
Всі клітини та внутрішньоклітинні органели оточені мембранами, які відіграють важливу роль у їх структурній організації та функціонуванні. Основні засади побудови всіх мембран однакові. Однак плазматична мембрана, а також мембрани ендоплазматичного ретикулума, апарату Гольджі, мітохондрій та ядра мають суттєві структурні особливості, вони унікальні за своїм складом та за характером виконуваних функцій.
Мембрани:
• відокремлюють клітини від довкілля та поділяють її на компартменти (відсіки);
• регулюють транспорт речовин у клітини та органели та у зворотному напрямку;
• забезпечують специфіку міжклітинних контактів;
• сприймають сигнали із зовнішнього середовища.
Узгоджене функціонування мембранних систем, що включають рецептори, ферменти, транспортні системи, допомагає підтримувати гомеостаз клітини та швидко реагувати на зміни стану довкілля шляхом регуляції метаболізму всередині клітин.
Біологічні мембрани побудовані з ліпідів та білків, пов'язаних один з одним за допомогою нековалентнихвзаємодій. Основу мембрани складає подвійний ліпідний шар,до складу якого включено білкові молекули (рис. 4.1). Ліпідний бислой утворений двома рядами амфіфільнихмолекул, гідрофобні «хвости» яких заховані всередину, а гідрофільні групи – полярні «головки» звернені назовні та контактують із водним середовищем.
1. Ліпіди мембран.До складу ліпідів мембран входять як насичені, і ненасичені жирні кислоти. Ненасичені жирні кислоти зустрічаються вдвічі частіше, ніж насичені, що визначає плинністьмембран та конформаційну лабільність мембранних білків.
У мембранах присутні ліпіди трьох головних типів – фосфоліпіди, гліколіпіди та холестерол (рис. 4.2 – 4.4). Найчастіше зустрічаються гліцерофосфоліпіди – похідні фосфатидної кислоти.
Рис. 4.1. Поперечний розріз плазматичної мембрани
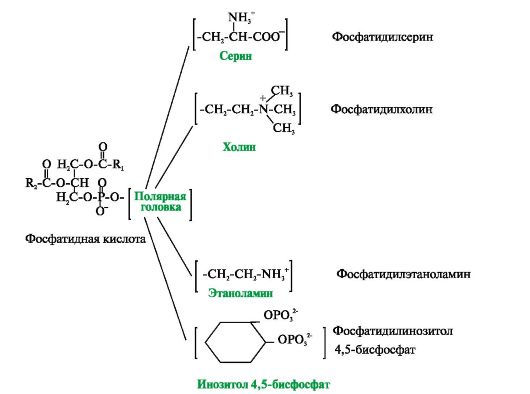 Рис. 4.2. Гліцерофосфоліпіди.
Рис. 4.2. Гліцерофосфоліпіди.
Фосфатидна кислота – це діацилгліцеролфосфат. R 1 , R 2 - радикали жирних кислот (гідрофобні "хвости"). З другим вуглецевим атомом гліцеролу пов'язаний залишок поліненасиченої жирної кислоти. Полярною «головкою» є залишок фосфорної кислоти та приєднана до нього гідрофільна група серину, холіну, етаноламіну або інозитолу
Існують також ліпіди – похідні аміноспирту сфінгозину.
Аміноспирт сфінгозин під час ацилювання, тобто. приєднання жирної кислоти до NH 2 -групи, перетворюється на церамід. Цераміди розрізняються по залишку жирної кислоти. З ОН-групою цераміду можуть бути пов'язані різні полярні групи. Залежно від будови полярної «головки» ці похідні поділені на дві групи – фосфоліпіди та гліколіпіди. Будова полярної групи сфінгофосфоліпідів (сфінгомієлінів) подібна до гліцерофосфоліпідів. Багато сфінгомієлінів міститься у складі мієлінових оболонок нервових волокон. Гліколіпіди є вуглеводні похідні цераміду. Залежно від будови вуглеводної складової розрізняють цереброзиди та гангліозиди.
Холестеролміститься в мембранах всіх тварин клітин, він надає мембранам жорсткості та знижує їх рідинність(плинність). Молекула холестеролу розташовується в гідрофобній зоні мембрани паралельно гідрофобним "хвостам" молекул фосфо-і гліколіпідів. Гідроксильна група холестеролу, як і гідрофільні «головки» фосфо- та гліколіпідів,
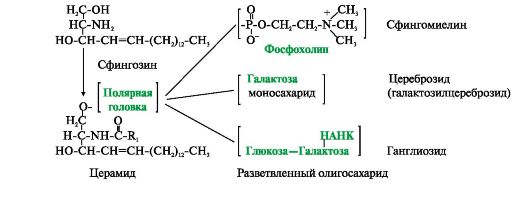 Рис. 4.3. Похідні аміноспирту сфінгозину.
Рис. 4.3. Похідні аміноспирту сфінгозину.
Церамід - ацильований сфінгозин (R 1 - радикал жирної кислоти). До фосфоліпідів належать сфінгомієліни, у яких полярна група складається з залишку фосфорної кислоти та холіну, етаноламіну або серину. Гідрофільною групою (полярною «головкою») гліколіпідів є вуглеводний залишок. Цереброзиди містять моноілі олігосахаридний залишок лінійної будови. До складу гангліозидів входить розгалужений олігосахарид, однією з мономерних ланок якого є НАНК - N-ацетилнейрамінова кислота
звернена до водної фази. Молярне співвідношення холестеролу та інших ліпідів у мембранах дорівнює 0,3-0,9. Найвище значення має ця величина для цитоплазматичної мембрани.
Збільшення вмісту холестеролу в мембранах зменшує рухливість ланцюгів жирних кислот, що впливає на конформаційну лабільність мембранних білків та знижує можливість їх латеральної дифузії.При підвищенні плинності мембран, спричинених дією на них ліпофільних речовин або перекисним окисленням ліпідів, частка холестеролу в мембранах зростає.
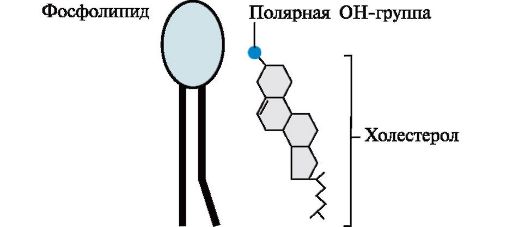 Рис. 4.4. Положення в мембрані фосфоліпідів та холестеролу.
Рис. 4.4. Положення в мембрані фосфоліпідів та холестеролу.
Молекула холестеролу складається з жорсткого гідрофобного ядра та гнучкого вуглеводневого ланцюга. Полярною «головкою» є ОН-група у 3-го вуглецевого атома молекули холестеролу. Для порівняння малюнку представлено схематичне зображення фосфоліпіду мембран. Полярна головка цих молекул значно більша і має заряд.
Ліпідний склад мембран різний, вміст того чи іншого ліпіду, мабуть, визначається різноманітністю функцій, які виконують ці молекули у мембранах.
Основні функції ліпідів мембран полягають у тому, що вони:
• формують ліпідний бислой – структурну основу мембран;
• забезпечують необхідне для функціонування мембранних білків середовище;
• беруть участь у регуляції активності ферментів;
• служать "якорем" для поверхневих білків;
• беруть участь у передачі гормональних сигналів.
Зміна структури ліпідного бісла може призвести до порушення функцій мембран.
2. Білки мембран.Білки мембран розрізняються за своїм становищем у мембрані (рис. 4.5). Мембранні білки, що контактують з гідрофобною областю ліпідного бислоя, мають бути амфіфільними, тобто. мати неполярний домен. Амфіфільність досягається завдяки тому, що:
• амінокислотні залишки, що контактують з ліпідним бислоем, переважно неполярні;
• багато мембранних білків ковалентно пов'язані з залишками жирних кислот (ацильовані).
Ацильні залишки жирних кислот, приєднані до білка, забезпечують його «заякорення» в мембрані та можливість латеральної дифузії. Крім того, білки мембран піддаються таким посттрансляційним модифікаціям, як глікозилювання та фосфорилювання. Глікозилювання зовнішньої поверхні інтегральних білків захищає їхню відмінність від ушкодження протеазами міжклітинного простору.
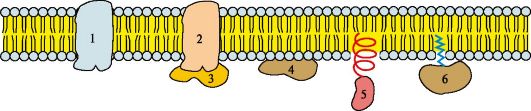 Рис. 4.5. Білки мембран:
Рис. 4.5. Білки мембран:
1, 2 - інтегральні (трансмембранні) білки; 3, 4, 5, 6 – поверхневі білки. В інтегральних білках частина поліпептидного ланцюга занурена в ліпідний шар. Ті ділянки білка, які взаємодіють із вуглеводневими ланцюгами жирних кислот, містять переважно неполярні амінокислоти. Ділянки білка, що знаходяться в ділянці полярних «головок», збагачені гідрофільними амінокислотними залишками. Поверхневі білки у різний спосіб прикріплюються до мембрани: 3 - пов'язані з інтегральними білками; 4 - приєднані до полярних «головок» ліпідного шару; 5 - «заякорені» у мембрані за допомогою короткого гідрофобного кінцевого домену; 6 - «заякорені» в мембрані за допомогою ковалентно зв'язаного ацильного залишку
Зовнішній та внутрішній шари однієї і тієї ж мембрани розрізняються за складом ліпідів та білків. Ця особливість у будові мембран називається трансмембранною асиметрією.
Білки мембран можуть брати участь у:
• вибірковий транспорт речовин у клітину та з клітини;
• передачу гормональних сигналів;
• утворенні «окаймлених ямок», що беруть участь в ендоцитозі та екзоцитозі;
• імунологічні реакції;
• якість ферментів у перетвореннях речовин;
• організації міжклітинних контактів, які забезпечують утворення тканин та органів.
ТЕМА 4.2. ТРАНСПОРТ РЕЧОВИН ЧЕРЕЗ МЕМБРАНИ
Одна з головних функцій мембран - регуляція перенесення речовин у клітину та з клітини, утримання речовин, які потрібні клітині та виведення непотрібних. Транспорт іонів, органічних молекул через мембрани може проходити по градієнту концентрації - пасивний транспорті проти градієнта концентрації - активний транспорт.
1. Пасивний транспортможе здійснюватися такими способами (рис. 4.6, 4.7):
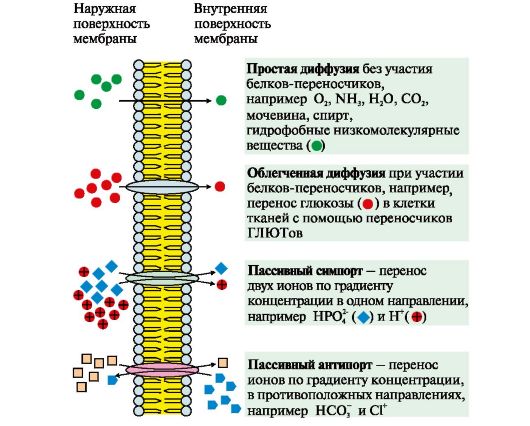 Рис. 4.6. Механізми перенесення речовин через мембрани за градієнтом концентрації
Рис. 4.6. Механізми перенесення речовин через мембрани за градієнтом концентрації
До пасивного транспорту належить дифузія іонів по білкових каналах,наприклад, дифузія Н+, Са 2 +, N+, К+. Функціонування більшості каналів регулюється специфічними лігандами чи зміною трансмембранного потенціалу.
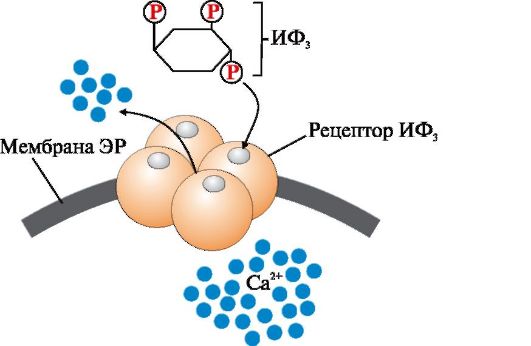 Рис. 4.7. Са 2 +-канал мембрани ендоплазматичного ретикулума, регульований інозитол-1,4,5-трифосфатом (ІФ 3).
Рис. 4.7. Са 2 +-канал мембрани ендоплазматичного ретикулума, регульований інозитол-1,4,5-трифосфатом (ІФ 3).
ІФ 3 (інозитол-1,4,5-трифосфат) утворюється при гідролізі мембранного ліпіду ФІФ 2 (фосфатидилінозитол-4,5-бісфосфату) під дією ферменту фосфоліпази С. ІФ 3 зв'язується специфічними центрами протомерів Са 2+- каналу. Змінюється конформація білка і канал відкривається - Са 2+ надходить у цитозоль клітини за градієнтом концентрації
2. Активний транспорт. Первинно-активнийтранспорт відбувається проти градієнта концентрації з витратою енергії АТФ за участю транспортних АТФаз, наприклад, Na+, К+-АТФаза, Н+-АТФаза, Са 2 +-АТФаза (рис. 4.8). Н+-АТФази функціонують як протонні насоси, за допомогою яких створюється кисле середовище у лізосомах клітини. За допомогою Са 2+ -АТФази цитоплазматичної мембрани та мембрани ендоплазматичного ретикулуму підтримується низька концентрація кальцію в цитозолі клітини та створюється внутрішньоклітинне депо Са 2+ у мітохондріях та ендоплазматичному ретикулумі.
Вторинно-активнийтранспорт відбувається за рахунок градієнта концентрації однієї з речовин, що переносяться (рис. 4.9), який створюється найчастіше Na+, К+-АТФазою, що функціонує з витратою АТФ.
Приєднання до активного центру білка-переносника речовини, концентрація якого вище, змінює його конформацію та збільшує спорідненість до сполуки, яка проходить у клітину проти концентрації градієнта. Вторинно-активний транспорт буває двох типів: активний сімпорті антипорт.
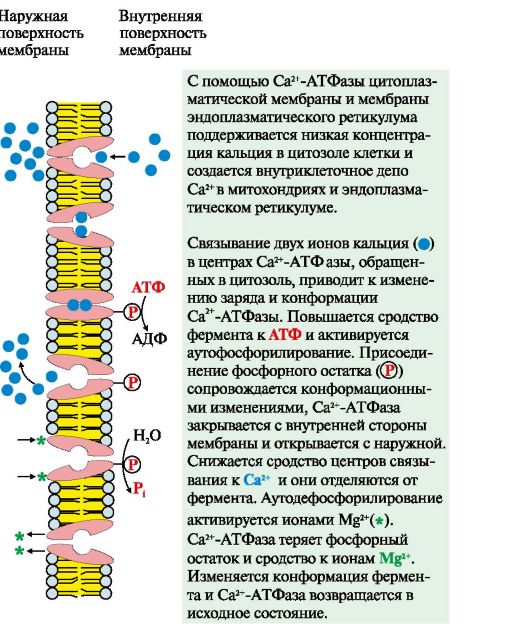 Рис. 4.8. Механізм функціонування Са 2 +-АТФази
Рис. 4.8. Механізм функціонування Са 2 +-АТФази
 Рис. 4.9. Вторинно-активний транспорт
Рис. 4.9. Вторинно-активний транспорт
3. Перенесення макромолекул та частинок за участю мембран – ендоцитоз та екзоцитоз.
Перенесення із позаклітинного середовища в клітину макромолекул, наприклад білків, нуклеїнових кислот, полісахаридів або ще більших частинок відбувається шляхом ендоцитозу.Зв'язування речовин або високомолекулярних комплексів відбувається у певних ділянках плазматичної мембрани, які називаються облямованими ямками.Ендоцитоз, що відбувається за участю рецепторів, вбудованих в облямовані ямки, дозволяє клітинам поглинати специфічні речовини і називається рецептор-залежним ендоцитозом.
Макромолекули, наприклад пептидні гормони, травні ферменти, білки позаклітинного матриксу, ліпопротеїнові комплекси секретуються в кров або міжклітинний простір шляхом екзоцитозу.Цей спосіб транспорту дозволяє виводити із клітини речовини, які накопичуються в секреторних гранулах. Найчастіше екзоцитоз регулюється шляхом зміни концентрації іонів кальцію в цитоплазмі клітин.
ТЕМА 4.3. ТРАНСМЕМБРАННА ПЕРЕДАЧА СИГНАЛІВ
Важлива властивість мембран - здатність сприймати та передавати всередину клітини сигнали з навколишнього середовища. Сприйняття клітинами зовнішніх сигналів відбувається за їх взаємодії з рецепторами, які у мембрані клітин-мішеней. Рецептори, приєднуючи сигнальну молекулу, активують внутрішньоклітинні шляхи передачі, це призводить до зміни швидкості різних метаболічних процесів.
1. Сигнальна молекула,специфічно взаємодіє з мембранним рецептором, називається первинним месенджером.Як первинні месенджери виступають різні хімічні сполуки - гормони, нейромедіатори, ейкозаноїди, ростові фактори або фізичні фактори, наприклад квант світла. Рецептори клітинної мембрани, активовані первинними месенджерами, передають отриману інформацію системі білків та ферментів, що утворюють каскад передачі сигналу,забезпечує посилення сигналу в кілька сотень разів. Час відповіді клітини, що полягає в активації або інактивації метаболічних процесів, скорочення м'язів, транспорту речовин з клітин-мішеней, може становити кілька хвилин.
Мембранні рецепториподіляються на:
• рецептори, що містять субодиницю, що зв'язує первинний месенджер, та іонний канал;
• рецептори, здатні виявляти каталітичну активність;
• рецептори, які за допомогою G-білків активують утворення вторинних (внутрішньоклітинних) месенджерів, що передають сигнал специфічним білкам та ферментам цитозолю (рис. 4.10).
Побічні месенджери мають невелику молекулярну масу, з високою швидкістю дифундують у цитозолі клітини, змінюють активність відповідних білків, а потім швидко розщеплюються або видаляються з цитозолю.
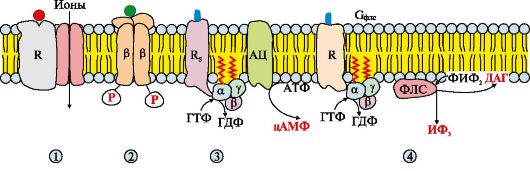 Рис. 4.10. Рецептори, локалізовані у мембрані.
Рис. 4.10. Рецептори, локалізовані у мембрані.
Мембранні рецептори можна поділити на три групи. Рецептори: 1 - містять субодиницю, що зв'язує сигнальну молекулу та іонний канал, наприклад, рецептор ацетилхоліну на постсинаптичній мембрані; 2 - виявляють каталітичну активність після приєднання сигнальної молекули, наприклад, рецептор інсуліну; 3, 4 - сигнали, що передають на фермент аденілатциклазу (АЦ) або фосфоліпазу С (ФЛС) за участю мембранних G-білків, наприклад різні типи рецепторів адреналіну, ацетилхоліну та інших сигнальних молекул
Роль вторинних месенджеріввиконують молекули та іони:
• цАМФ (циклічний аденозин-3",5"-монофосфат);
• цГМФ (циклічний гуанозин-3",5"-монофосфат);
• ІФ 3 (інозитол-1,4,5-трифосфат);
• ДАГ (діацилгліцерол);
Існують гормони (стероїдні та тиреоїдні), які, проходячи ліпідний бислой, проникають у клітинуі взаємодіють з внутрішньоклітинними рецепторами.Фізіологічно важливою різницею між мембранними і внутрішньоклітинними рецепторами є швидкість відповіді на сигнал, що надходить. У першому випадку ефект буде швидким та нетривалим, у другому – повільним, але тривалим.
Рецептори, пов'язані з G-білками
Взаємодія гормонів з рецепторами, пов'язаними з G-білками, призводить до активації інозитолфосфатної системи трансдукції сигналу або зміни активності аденілатциклазної регуляторної системи.
2. Аденілатциклазна системавключає (рис. 4.11):
- інтегральнібілки цитоплазматичної мембрани:
• R s – рецептор первинного месенджера – активатора аденілатциклазної системи (АЦС);
• R; - рецептор первинного месенджера – інгібітора АЦС;
• фермент аденілатциклаз (АЦ).
- «заякорені»білки:
• G s - ГТФ-зв'язуючий білок, що складається з α,βγ-субодиниць, в якому (α,-субодиниця пов'язана з молекулою ГДФ;
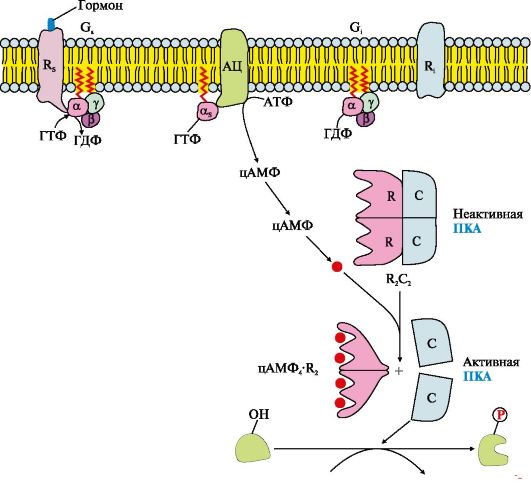
Рис. 4.11. Функціонування аденілатциклазної системи
• G; - ГТФ-зв'язуючий білок, що складається з αβγ-субодиниць, в якому а; -субодиниця пов'язана з молекулою ГДФ; - цитозольнийфермент протеїнкіназу А (ПКА).
Послідовність подій передачі сигналу первинних месенджерів за допомогою аденілатциклазної системи
Рецептор має центри зв'язування первинного месенджера на зовнішній поверхні мембрани та G-білка (α,βγ-ГДФ) на внутрішній поверхні мембрани. Взаємодія активатора аденілатциклазної системи, наприклад, гормону з рецептором (R s), призводить до зміни конформації рецептора. Збільшується спорідненість рецептора до G-білка. Приєднання комплексу гормон-рецептор до GS-ГДФ знижує спорідненість α-субодиниці G..-білка до ГДФ і збільшує спорідненість до ГТФ. В активному центрі α-субодиниці ГДФ заміщається на ГТФ. Це викликає зміну конформації субодиниці α і зниження її спорідненості до субодиниця βγ. Субодиниця α,-ГТФ, що відокремилася, латерально переміщається в ліпідному шарі мембрани до ферменту аденілатциклазі.
Взаємодія α,-ГТФ з регуляторним центром аденілатциклази змінює конформацію ферменту, призводить до його активації та збільшення швидкості утворення вторинного месенджера - циклічного аденозин-3",5"-монофосфату (цАМФ) з АТФ. У клітині підвищується концентрація цАМФ. Молекули цАМФ можуть оборотно з'єднуватися з регуляторними субодиницями протеїнкінази А (ПКА), яка складається з двох регуляторних (R) та двох каталітичних (С) субодиниць - (R 2 С 2). Комплекс R 2 З 2 ферментативної активністю не має. Приєднання цАМФ до регуляторних субодиниць викликає зміну їх конформації та втрату комплементарності до С-субодиниця. Каталітичні субодиниці набувають ферментативної активності.
Активна протеїнкіназа А за допомогою АТФ фосфорилює специфічні білки по залишках серину та треоніну. Фосфорилювання білків і ферментів підвищує або знижує їхню активність, тому змінюється швидкість метаболічних процесів, в яких вони беруть участь.
Активація сигнальною молекулою рецептора R стимулює функціонування Gj-білка, яке протікає за тими самими правилами, що і для G-білка. Але при взаємодії субодиниці αi-ГТФ з аденілатциклазою активність ферменту знижується.
Інактивація аденілатциклази та протеїнкінази А
α,-субодиниця в комплексі з ГТФ при взаємодії з аденілатциклазою починає проявляти ферментативну (ГТФ-фосфатазну) активність, вона гідролізує ГТФ. Утворена молекула ГДФ залишається в активному центрі α-субодиниці, змінює її конформацію і зменшує спорідненість до АЦ. Комплекс АЦ та α,-ГДФ дисоціює, α,-ГДФ включається до G..-білок. Відділення α-ГДФ від аденілатциклази інактивує фермент і синтез цАМФ припиняється.
Фосфодіестераза- «заякорений» фермент цитоплазматичної мембрани гідролізує молекули цАМФ, що утворилися раніше, до АМФ. Зниження концентрації цАМФ у клітині викликає розщеплення комплексу цАМФ 4 К" 2 і підвищує спорідненість R-і С-субодиниць, утворюється неактивна форма ПКА.
Фосфорильовані ферменти та білки під дією фосфопротеїнфосфатазипереходять у дефосфорильовану форму, змінюється їх конформація, активність та швидкість процесів, у яких беруть участь ці ферменти. В результаті система входить у вихідний стан і готова знову активуватися при взаємодії гормону з рецептором. Таким чином, забезпечується відповідність вмісту гормону в крові та інтенсивності відповіді клітин-мішеней.
3. Участь аденілатциклазної системи у регуляції експресії генів.Багато білкових гормонів: глюкагон, вазопресин, паратгормон та ін., що передають свій сигнал за допомогою аденілатциклазної системи, можуть не тільки викликати зміну швидкості реакцій шляхом фосфорилювання вже наявних у клітині ферментів, але і збільшувати або зменшувати їх кількість, регулюючи експресію генів (рис. 4.1). ). Активна протеїнкіназа А може проходити в ядро та фосфорилювати фактор транскрипції (СРЄВ). Приєднання фосфорного
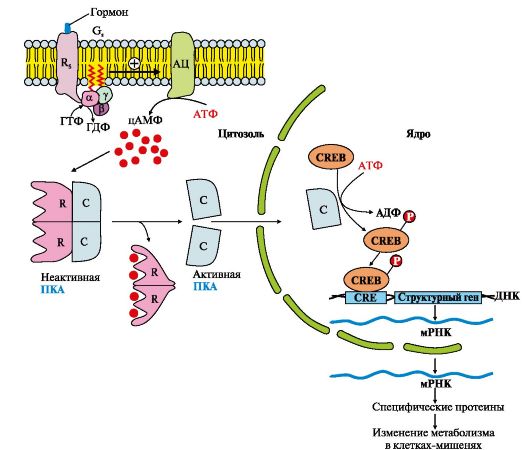 Рис. 4.12. Аденілатциклазний шлях, що призводить до експресії специфічних генів
Рис. 4.12. Аденілатциклазний шлях, що призводить до експресії специфічних генів
залишку підвищує спорідненість фактора транскрипції (СРЕВ-(Р) до специфічної послідовності регуляторної зони ДНК-СRЕ (цАМФ-response element) і стимулює експресію генів певних білків.
Синтезовані білки можуть бути ферментами, збільшення кількості яких підвищує швидкість реакцій метаболічних процесів, або мембранними переносниками, що забезпечують надходження або вихід з клітин певних іонів, води або інших речовин.

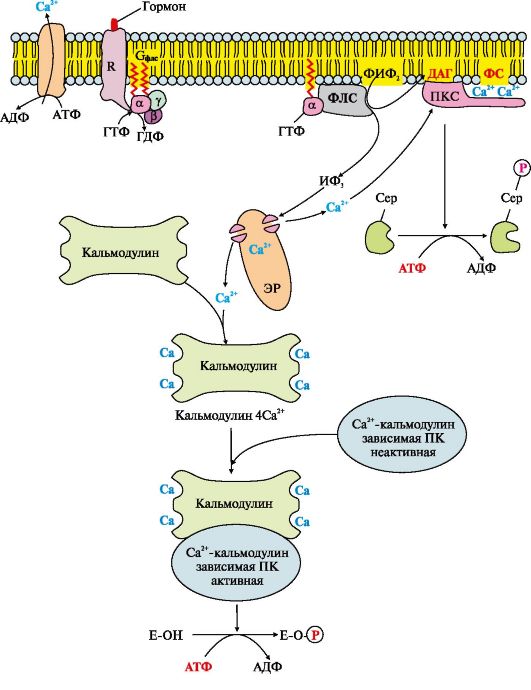 Рис. 4.13. Інозитолфосфатна система
Рис. 4.13. Інозитолфосфатна система
Роботу системи забезпечують білки: кальмодулін, фермент протеїнкіназа С, Са2+-кальмодулін-залежні протеїнкінази, регульовані Са2+-канали мембрани ендоплазматичного ретикулуму, Са2+-АТФази клітинної та мітохондріальної мембран.
Послідовність подій передачі сигналу первинних месенджерів за допомогою інозитолфосфатної системи
Зв'язування активатора інозитолфосфатної системи з рецептором (R) призводить до зміни його конформації. Підвищується спорідненість рецептора до Gф лс-білку. Приєднання комплексу первинний месенджер-рецептор до Gф лс-ГДФ знижує спорідненість аф лс-субодиниці до ГДФ і збільшує спорідненість до ГТФ. В активному центрі аф лс-субодиниці ГДФ заміщається на ГТФ. Це викликає зміну конформації субодиниці аф лс і зниження спорідненості до субодиницям βγ, відбувається дисоціація Gф лс -білка. Відокремлена субодиниця аф лс -ГТФ латерально переміщається мембраною до ферменту фосфоліпаз С.
Взаємодія аф лс -ГТФ з центром зв'язування фосфоліпази С змінює конформацію та активність ферменту, зростає швидкість гідролізу фосфоліпіду клітинної мембрани - фосфатидилінозитол-4,5-бісфосфату (ФІФ 2) (рис. 4.14).
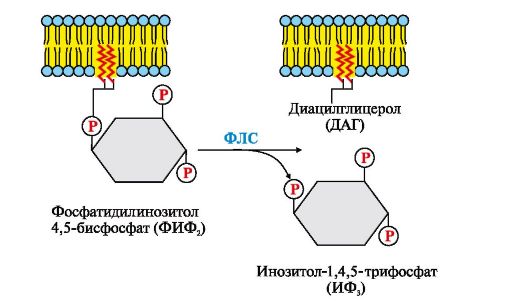 Рис. 4.14. Гідроліз фосфатидилінозитол-4,5-бісфосфату (ФІФ 2)
Рис. 4.14. Гідроліз фосфатидилінозитол-4,5-бісфосфату (ФІФ 2)
У ході реакції утворюються два продукти - вторинні вісники гормонального сигналу (вторинні месенджери): діацилгліцерол, який залишається в мембрані та бере участь в активації ферменту протеїнкінази С, та інозитол-1,4,5-трифосфат (ІФ 3), який є гідрофільною сполукою іде в цитозоль. Таким чином сигнал, прийнятий рецептором клітини, роздвоюється. ІФ 3 зв'язується специфічними центрами Са 2+ - каналу мембрани ендоплазматичного ретикулума (Е)), що призводить до зміни конформації білка та відкриття Са 2+ -каналу. Оскільки концентрація кальцію в ЕР приблизно на 3-4 порядки вища, ніж у цитозолі, після відкриття каналу Са 2+ за градієнтом концентрації надходить у цитозоль. У відсутність ІФ 3 у цитозолі канал закритий.
У цитозолі всіх клітин міститься невеликий білок кальмодулін, що має чотири центри зв'язування Са 2+ . При підвищенні концентрації
кальцію він активно приєднується до кальмодуліну, утворюючи комплекс 4Са 2+ -кальмодулін. Цей комплекс взаємодіє з Са 2+ -кальмодулінзалежними протеїнкіназами, іншими ферментами та підвищує їх активність. Активована Са2+-кальмодулін-залежна протеїнкіназа фосфорилює певні білки та ферменти, внаслідок чого змінюється їх активність та швидкість метаболічних процесів, у яких вони беруть участь.
Підвищення концентрації Са 2+ у цитозолі клітини збільшує швидкість взаємодії Са 2+ з неактивним цитозольним ферментом протеїнкіназою С (ПКС).Зв'язування ПКС з іонами кальцію стимулює переміщення білка до плазматичної мембрани та дозволяє ферменту вступати у взаємодію з негативно зарядженими «головками» молекул фосфатидилсерину (ФС) мембрани. Діацилгліцерол, займаючи специфічні центри в протеїнкіназі С, ще більшою мірою збільшує її спорідненість до іонів кальцію. На внутрішній стороні мембрани утворюється активна форма ПКС (ПКС? Са 2+? ФС? ДАГ), яка фосфорилює специфічні ферменти.
Включення ІФ-системи нетривало, і після відповіді клітини на стимул відбувається інактивація фосфоліпази С, протеїнкінази С і Са2+-кальмодулін-залежних ферментів. аф лс -Субодиниця в комплексі з ГТФ і фосфоліпазою С проявляє ферментативну (ГТФ-фосфатазну) активність, вона гідролізує ГТФ. Пов'язана з ГДФ аф лс -субодиниця втрачає спорідненість до фосфоліпазі З повертається у вихідне неактивне стан, тобто. включається в комплекс αβγ-ГДФ (Gф лс -білок).
Відділення аф лс-ГДФ від фосфоліпази С інактивує фермент і гідроліз ФІФ 2 припиняється. Підвищення концентрації Са 2+ у цитозолі активує роботу Са 2+ -АТФаз ендоплазматичного ретикулуму, цитоплазматичної мембрани, які «викачують» Са 2+ із цитозолю клітини. У цьому процесі беруть участь також Na+/Са 2+- та Н+/Са 2+-переносники, що функціонують за принципом активного антипорту. Зниження концентрації Са 2+ призводить до дисоціації та інактивації Са 2+ -кальмодулінзалежних ферментів, а також втрати спорідненості протеїнкінази С до ліпідів мембрани та зниження її активності.
ІФ 3 і ДАГ, що утворилися в результаті активації системи, можуть знову взаємодіяти один з одним і перетворюватися на фосфатидилінозитол-4,5-бісфосфат.
Фосфорильовані ферменти та білки під дією фосфопротеінфосфатази переходять у дефосфорильовану форму, змінюється їх конформація та активність.
5. Каталітичні рецептори.Каталітичні рецептори є ферментами. Активаторами цих ферментів може бути гормони, ростові чинники, цитокини. В активній формі - рецептори-ферменти фосфорилують специфічні білки по -ОН-груп тирозину, тому їх називають тирозиновими протеїнкіназами (рис. 4.15). За участю спеціальних механізмів сигнал, отриманий каталітичним рецептором, може бути переданий у ядро, де стимулює або пригнічує експресію певних генів.
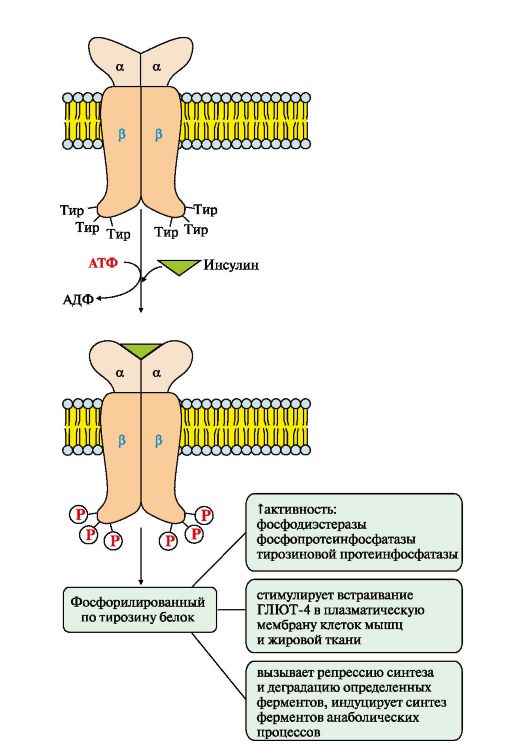 Рис. 4.15. Активація інсулінового рецептора.
Рис. 4.15. Активація інсулінового рецептора.
Фосфопротеїнфосфатаза дефосфорілює специфічні фосфопротеїни.
Фосфодіестераза перетворює цАМФ на АМФ і цГМФ на ГМФ.
ГЛЮТ 4 – переносники глюкози в інсулінзалежних тканинах.
Тирозинова протеїнфосфатаза дефосфорилює β-субодиниці рецептора
інсуліну
Прикладом каталітичного рецептора може бути рецептор інсуліну,до складу якого входять дві а-і дві β-субодиниці. а-субодиниці розташовані на зовнішній поверхні клітинної мембрани, β-субодиниці пронизують мембранний бислой. Центр зв'язування інсуліну утворений N-кінцевими доменами а-субодиниць. Каталітичний центр рецептора знаходиться на внутрішньоклітинних доменах β-субодиниць. Цитозольна частина рецептора має кілька залишків тирозину, які можуть бути фосфориловані та дефосфориловані.
Приєднання інсуліну до центру зв'язування, утворений а-субодиницями, викликає кооперативні конформаційні зміни рецептора. β-субодиниці виявляють тирозинкіназну активність і каталізують трансаутофосфорилювання (перша β-субодиниця фосфорилює другу β-субодиницю, і навпаки) по кількох залишках тирозину. Фосфорилювання призводить до зміни заряду, конформації та субстратної специфічності ферменту (Тир-ПК). Тирозинова-ПК фосфорилює певні клітинні білки, які отримали назву субстратів інсулінового рецептора. У свою чергу, ці білки беруть участь в активації каскаду реакцій фосфорилювання:
• фосфопротеїнфосфатази(ФПФ), яка дефосфорілює специфічні фосфопротеїни;
• фосфодіестерази,яка перетворює цАМФ на АМФ і цГМФ на ГМФ;
• ГЛЮТ 4- переносників глюкози в інсулінзалежних тканинах, тому підвищується надходження глюкози до клітин м'язів та жирової тканини;
• тирозинової протеїнфосфатази,яка дефосфорилює β-субодиниці рецептора інсуліну;
• регуляторних білків ядра, факторів транскрипції,генів, що підвищують або знижують експресію, певних ферментів.
Реалізація ефекту ростових факторівможе здійснюватися за допомогою каталітичних рецепторів, які складаються з одного поліпептидного ланцюга, але при зв'язуванні первинного месенджера утворюють димери. Всі рецептори цього типу мають позаклітинний глікозильований домен, трансмембранний (аспіраль) і цитоплазматичний домен, здатний при активації проявляти протеїнкіназну активність.
Димеризація сприяє активації їх каталітичних внутрішньоклітинних доменів, які здійснюють трансаутофосфорилювання по амінокислотних залишках серину, треоніну або тирозину. Приєднання фосфорних залишків призводить до формування у рецептора центрів зв'язування для специфічних цитозольних білків та активації протеїнкіназного каскаду передачі сигналу (рис. 4.16).
Послідовність подій передачі сигналу первинних месенджерів (ростових факторів) за участю Ras- та Raf-білків.
Зв'язування рецептора (R) з фактором росту (ФР) призводить до його димеризації та трансаутофосфорилування. Фосфорильований рецептор набуває спорідненості з Grb2-білком. Утворений комплекс ФР*R*Grb2 взаємодіє із цитозольним білком SOS. Зміна конформації SOS
забезпечує його взаємодію із заякореним білком мембрани Ras-ГДФ. Утворення комплексу ФР?R?Gгb2?SOS?Ras-ГДФ знижує спорідненість Ras-білка до ГДФ і збільшує спорідненість до ГТФ.
Заміна ГДФ на ГТФ змінює конформацію Ras-білка, який відокремлюється від комплексу та взаємодіє з Raf-білком у примембранній області. Комплекс Ras-ГТФ-Raf проявляє протеїнкіназну активність і фосфорилює фермент МЕК-кіназу. Активована МЕК-кіназа у свою чергу фосфорилює МАП-кіназу по треоніну та тирозину.
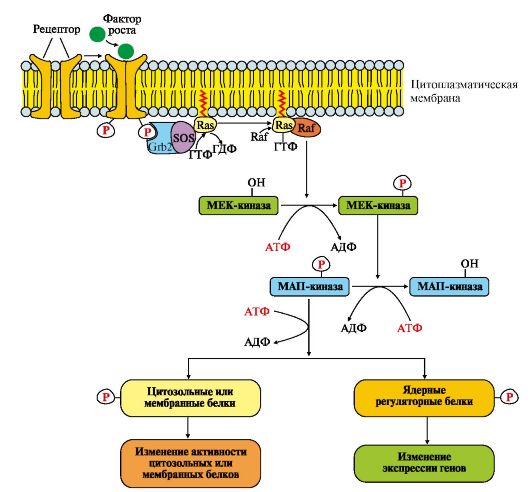 Рис.4.16. МАП-кіназний каскад.
Рис.4.16. МАП-кіназний каскад.
Рецептори такого типу мають епідермальний фактор росту (ЕФР), фактор росту нервів (ФРН) та інші фактори росту.
Grb2 - протеїн, що взаємодіє з рецептором ростового фактора (growth receptor binding protein); SOS (GEF) - ГДФ-ГТФ обмінний фактор (guanine nucleotide exchange factor); Ras – G-білок (гуанідинтрифосфатаза); Raf-кіназа - в активній формі - фосфорилююча МЕК-кіназу; МЕК-кіназа – кіназа МАП-кінази; МАП-кіназа – мітогенактивована протеїнкіназа (mitogen-aktivated protein kinase)
Приєднання групи -РО 3 2 - до амінокислотних радикалів МАП-кінази змінює її заряд, конформацію та активність. Фермент фосфорилює по серину та треоніну специфічні білки мембран, цитозолю та ядра.
Зміна активності цих білків впливає на швидкість метаболічних процесів, функціонування мембранних транслоказів, мітотичну активність клітин-мішеней.
Рецептори з гуанілатциклазною активністютакож відносяться до каталітичних рецепторів. Гуанілатциклазакаталізує утворення з ГТФ цГМФ, який є одним із важливих месенджерів (посередників) внутрішньоклітинної передачі сигналу (рис. 4.17).
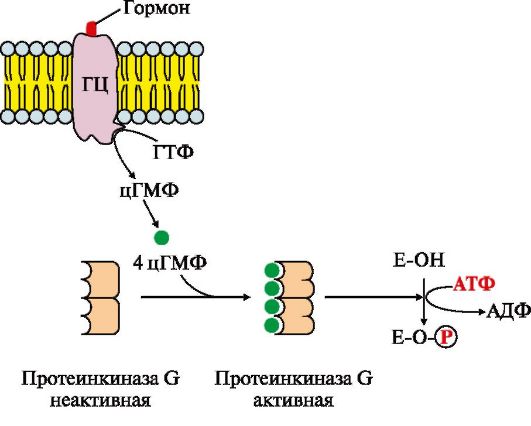 Рис. 4.17. Регулювання активності мембранної гуанілатциклази.
Рис. 4.17. Регулювання активності мембранної гуанілатциклази.
Мембранно-пов'язана гуанілатциклаза (ГЦ) – трансмембранний глікопротеїн. Центр зв'язування сигнальної молекули знаходиться на позаклітинному домені, внутрішньоклітинний домен гуанілатциклази внаслідок активації виявляє каталітичну активність
Приєднання первинного месенджера до рецептора активує гуанілатциклазу, яка каталізує перетворення ГТФ на циклічний гуанозин-3",5"-монофосфат (цГМФ) - вторинний месенджер. У клітині підвищується концентрація цГМФ. Молекули цГМФ можуть оборотно приєднуватися до регуляторних центрів протеїнкінази G (ПКЧ5), що складається з двох субодиниць. Чотири молекули цГМФ змінюють конформацію та активність ферменту. Активна протеїнкіназа G каталізує фосфорилювання певних білків та ферментів цитозолю клітини. Одним із первинних месенджерів протеїнкінази G є передсердний натріуретичний фактор (ПНФ), що регулює гомеостаз рідини в організмі.
6. Передача сигналу за допомогою внутрішньоклітинних рецепторів.Гідрофобні за хімічною природою гормони (стероїдні гормони та тироксин) можуть дифундувати через мембрани, тому їх рецептори знаходяться у цитозолі або ядрі клітини.
Цитозольні рецептори пов'язані з білком-шапероном, який запобігає передчасній активації рецептора. Ядерні та цитозольні рецептори стероїдних та тиреоїдних гормонів містять ДНК-зв'язуючий домен, що забезпечує в ядрі взаємодію комплексу гормон-рецептор з регуляторними ділянками ДНК та зміну швидкості транскрипції.
Послідовність подій, що призводять до зміни швидкості транскрипції
Гормон проходить через подвійний ліпідний шар клітинної мембрани. У цитозолі чи ядрі гормон взаємодіє з рецептором. Комплекс гормон-рецептор проходить в ядро і приєднується до регуляторної нуклеотидної послідовності ДНК. енхансеру(рис. 4.18) або сайленсер.Доступність промотору для РНК-полімерази збільшується при взаємодії з енхансером або зменшується при взаємодії із сайленсером. Відповідно збільшується чи зменшується швидкість транскрипції певних структурних генів. Зрілі мРНК виходять із ядра. Збільшується чи зменшується швидкість трансляції певних білків. Змінюється кількість білків, які впливають на метаболізм та функціональний стан клітини.
У кожній клітині існують рецептори, включені до складу різних сигнал-трансдукторних систем, що перетворюють всі зовнішні сигнали у внутрішньоклітинні. Число рецепторів для конкретного первинного месенджера може варіювати в межах від 500 до 100 000 на клітину. Вони розміщуються на мембрані віддалено один від одного або зосереджені у певних її ділянках.
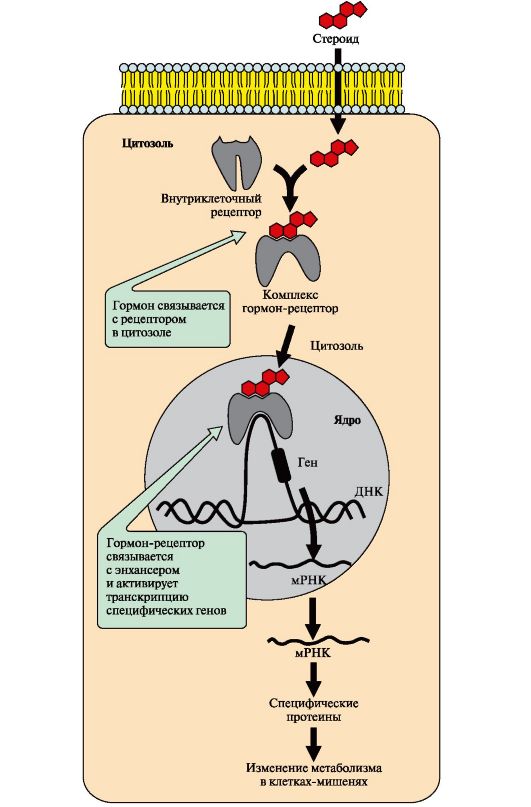 Рис. 4.18. Передача сигналу на внутрішньоклітинні рецептори
Рис. 4.18. Передача сигналу на внутрішньоклітинні рецептори
б) з таблиці виберіть ліпіди, які беруть участь у:
1. Активації протеїнкінази С
2. Реакції утворення ДАГ під дією фосфоліпази С
3. Формуванні мієлінових оболонок нервових волокон
в) напишіть реакцію гідролізу ліпіду, обраного вами у п. 2;
г) вкажіть, який із продуктів гідролізу бере участь у регуляції Са 2 +-каналу ендоплазматичного ретикулуму.
2. Виберіть правильні відповіді.
На конформаційну лабільність білків-переносників може впливати:
Б. Зміна електричного потенціалу на мембрані
B. Приєднання специфічних молекул Г. Жирнокислотний склад ліпідів бислоя Д. Кількість речовини, що переноситься
3. Встановіть відповідність:
A. Кальцієвий канал ЕР Б. Са 2 +-АТФаза
Г. Ка+-залежний переносник Са 2 + Д. N+, К+-АТФаза
1. Переносить Na+ за градієнтом концентрації
2. Функціонує за механізмом полегшеної дифузії
3. Переносить Na+ проти градієнта концентрації
4. Перенесіть табл. 4.2. у зошит та заповніть її.
Таблиця 4.2. Аденілатциклазна та інозитолфосфатна системи
Будова та етапи функціонування | Аденілатциклазна система | Інозитолфосфатна система |
Приклад первинного месенджера системи | ||
Інтегральний білок клітинної мембрани, що взаємодіє комплементарно з первинним месенджером | ||
Білок, що активує фермент сигнальної системи | ||
Фермент системи, що утворює вторинний(е) месенджер(и) | ||
Вторинний (і) месенджер (-и) системи | ||
Цитозольний (е) фермент (и) системи, що взаємодіє (е) з вторинним месенджером | ||
Механізм регуляції (у цій системі) активності ферментів метаболічних шляхів | ||
Механізми зниження концентрації вторинних месенджерів у клітині-мішені | ||
Причина зниження активності мембранного ферменту сигнальної системи |
ЗАВДАННЯ ДЛЯ САМОКОНТРОЛЮ
1. Встановіть відповідність:
A. Пасивний симпорт Б. Пасивний антипорт
B. Ендоцитоз Г. Екзоцитоз
Д. Первинно-активний транспорт
1. Транспорт речовини у клітину відбувається разом із частиною плазматичної мембрани
2. Одночасно в клітину по градієнту концентрації проходять дві різні речовини
3. Перенесення речовин іде проти градієнта концентрації
2. Виберіть правильну відповідь.
ag-Субодиниця G-білка, пов'язана з ГТФ, активує:
A. Рецептор
Б. Протеїнкіназу А
B. Фосфодіестеразу Г. Аденілатциклазу Д. Протеїнкіназу С
3. Встановіть відповідність.
Функція:
A. Регулює активність каталітичного рецептора Б. Активує фосфоліпазу
B. Переводить в активну форму протеїнкіназу А
Г. Підвищує концентрацію Са 2+ у цитозолі клітини Д. Активує протеїнкіназу С
Вторинний месенджер:
4. Встановіть відповідність.
Функціонування:
A. Здатний до латеральної дифузії в бислое мембрани
Б. У комплексі з первинним месенджером приєднується до енхансеру
B. Виявляє ферментативну активність при взаємодії з первинним месенджером
Г. Може взаємодіяти з G-білком
Д. У процесі передачі сигналу взаємодіє з фосфоліпазою Рецептор:
1. Інсуліну
2. Адреналіну
3. Стероїдного гормону
5. Виконайте «ланцюгове» завдання:
а) пептидні гормони взаємодіють з рецепторами:
A. У цитозолі клітини
Б. Інтегральними білками мембран клітин-мішеней
B. У ядрі клітини
Г. Ковалентно пов'язаними з ФІФ 2
б) взаємодія такого рецептора з гормоном викликає підвищення концентрації у клітині:
A. Гормона
Б. Проміжних метаболітів
B. Вторинних месенджерів Г. Ядерних білків
в) цими молекулами можуть бути:
A. ТАГ Б. ГТФ
B. ФІФ 2 Г. цАМФ
г) вони активують:
A. Аденілатциклазу
Б. Са 2+ -залежний кальмодулін
B. Протеїнкіназу А Г. Фосфоліпазу С
д) цей фермент змінює швидкість метаболічних процесів у клітині шляхом:
A. Підвищення концентрації Са 2+ у цитозолі Б. Фосфорилювання регуляторних ферментів
B. Активації протенфосфатази
Г. Зміни експресії генів регуляторних білків
6. Виконайте «ланцюгове» завдання:
а) приєднання фактора росту (ФР) до рецептора (R) призводить до:
A. Зміни локалізації комплексу ФР-R
Б. Димеризації та трансаутофосфорилювання рецептора
B. Зміни конформації рецептора та приєднання до Gs-білка Г. Переміщення комплексу ФР-R
б) такі зміни в структурі рецептора збільшують його спорідненість до поверхневого білка мембрани:
Б. Raf Г. Grb2
в) ця взаємодія підвищує ймовірність приєднання до комплексу цитозольного білка:
А. Кальмодуліну B. Ras
Б. ПКС Г. SOS
г) який збільшує комплементарність комплексу до «заякореного» білка:
д) зміна конформації «заякореного» білка знижує його спорідненість до:
А. ЦАМФ B. ГТФ
Б. ГДФ Г. АТФ
е) ця речовина замінюється на:
А. ГДФ B. АМФ
Б. ЦГМФ Г. ГТФ
ж) приєднання нуклеотиду сприяє взаємодії «заякореного» білка з:
А. ПКА B. Кальмодуліном
з) цей білок входить до складу комплексу, який фосфорилює:
А. МЕК-кіназу В. Протеїнкіназу С
Б. Протеїнкіназу А Г. МАП-кіназу
і) цей фермент у свою чергу активує:
А. МЕК-кіназу В. Протеїнкіназу G
Б. Raf-білок Г. МАП-кіназу
к) фосфорилювання білка підвищує його спорідненість до:
А. Білкам SOS та Raf В. Регуляторним білкам ядра Б. Кальмодуліну Г. Ядерним рецепторам
л) активація цих білків призводить до:
A. Дефосфорилування ГТФ в активному центрі білка Ras Б. Зниження спорідненості рецептора до фактору зростання
B. Підвищення швидкості матричних біосинтезів Г. Дисоціації комплексу SOS-Grb2
м) внаслідок цього:
A. Білок SOS відокремлюється від рецептора
Б. Відбувається дисоціація протомерів рецептора (R)
B. Ras-білок відокремлюється від Raf-білка
Г. Зростає проліферативна активність клітини-мішені.
ЕТАЛОНИ ВІДПОВІДЕЙ ДО «ЗАВАННЯМ ДЛЯ САМОКОНТРОЛЮ»
1. 1-В, 2-А, 3-Д
3. 1-В, 2-Д, 3-Г
4. 1-В, 2-Г, 3-Б
5. а) Б, б) В, в) Р, г) В, д) Б
6. а) Б, б) Р, в) Р, г) А, д) Б, е) Р, ж) Р, з) А, і) Р, к) В, л) В, м) Р
ОСНОВНІ ТЕРМІНИ І ПОНЯТТЯ
1. Структура та функції мембран
2. Транспорт речовин через мембрани
3. Особливості будови білків мембран
4. Трансмембранні системи передачі сигналів (аденілатциклазна, інозитолфосфатна, гуанілатциклазна, каталітичні та внутрішньоклітинні рецептори)
5. Первинні месенджери
6. Вторинні месенджери (посередники)
ЗАВДАННЯ ДЛЯ АУДИТОРНОЇ РОБОТИ
1. Ознайомтеся з рис. 4.19 та виконайте такі завдання:
а) назвіть вид транспорту;
б) встановіть порядок подій:
A. Cl - за градієнтом концентрації виходить із клітини
Б. Протеїнкіназа А фосфорилює R-субодиницю каналу
B. Змінюється конформація R-субодиниці
Г. Відбуваються кооперативні конформаційні зміни мембранного білка
Д. Активується аденілатциклазна система
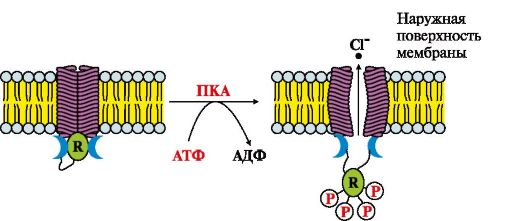 Рис. 4.19. Функціонування С1-каналу ендотелію кишечника.
Рис. 4.19. Функціонування С1-каналу ендотелію кишечника.
R - регуляторний білок, який переходить у фосфорильовану форму під дією протеїнкінази А (ПКА)
в) порівняйте функціонування Са 2+ -каналу мембрани ендоплазматичного ретикулуму та Cl - -каналу клітини ендотелію кишечника, заповнивши табл. 4.3.
Таблиця 4.3. Способи регулювання функціонування каналів
Розв'яжіть завдання
1. Скорочення серцевого м'яза активує Са 2+, вміст якого у цитозолі клітини підвищується за рахунок функціонування цАМФ-залежних переносників цитоплазматичної мембрани. У свою чергу, концентрація цАМФ у клітинах регулюється двома сигнальними молекулами – адреналіном та ацетилхоліном. Причому відомо, що адреналін, взаємодіючи з β 2 -адренорецепторами, підвищує концентрацію цАМФ у клітинах міокарда та стимулює серцевий викид, а ацетилхолін, взаємодіючи з М 2 -холінорецепторами, знижує рівень цАМФ та скоротливість міокарда. Поясніть, чому два первинних месенджери, використовуючи ту саму систему трансдукції сигналу, викликають різну клітинну відповідь. Для цього:
а) подайте схему передачі сигналу для адреналіну та ацетилхоліну;
б) вкажіть різницю в каскадах передачі сигналів цих месенджерів.
2. Ацетилхолін, взаємодіючи з М 3 -холінорецепторами слинних залоз, стимулює вихід Са 2+ з ЕР. Підвищення концентрації Са 2+ у цитозолі забезпечує екзоцитоз секреторних гранул і вивільнення в слинну протоку електролітів та невеликої кількості білків. Поясніть, як регулюється робота Са 2+ -каналів ЕР. Для цього:
а) назвіть вторинний месенджер, який забезпечує відкриття Са 2+ -каналів ЕР;
б) напишіть реакцію утворення вторинного месенджера;
в) представте схему трансмембранної передачі сигналу ацетилхоліну, в ході активації якої утворюється регуляторний ліганд Са 2+ -кана-
3. Дослідники інсулінового рецептора встановили значну зміну в гені білка - одного з субстратів інсулінового рецептора. Як порушення структурі цього білка позначиться на функціонуванні системи передачі сигналу інсуліну? Для відповіді на запитання:
а) наведіть схему трансмембранної передачі інсуліну сигналу;
б) назвіть білки та ферменти, які активує інсулін у клітинахмішенях, вкажіть їх функцію.
4. Білок Ras є "заякореним" білком цитоплазматичної мембрани. Функцію «якоря» виконує 15-вуглецевий залишок фарнезила Н 3 С-(СН 3)С=СН-СН 2 -[СН 2 -(СН 3)С=СН-СН 2 ] 2 -, який приєднується до білка ферментом фарнезилтрансферазою ході посттрансляційної модифікації. Нині інгібітори цього ферменту проходять клінічні випробування.
Чому використання цих препаратів призводить до порушення трансдукції сигналу ростових факторів? Для відповіді:
а) уявіть схему передачі сигналу за участю Ras-білків;
б) поясніть функцію Ras-білків та наслідки порушення їх ацилювання;
Природа створила безліч організмів і клітин, але, незважаючи на це, будова та більшість функцій біологічних мембран однакові, що дозволяє розглядати їх структуру та вивчати їх ключові властивості без прихильності до конкретного виду клітин.
Що таке мембрана?
Мембрани – це захисний елемент, який є невід'ємною складовою клітини будь-якого живого організму.
Структурною та функціональною одиницею всіх живих організмів на планеті є клітина. Життєдіяльність її нерозривно пов'язана з довкіллям, з яким вона обмінюється енергією, інформацією, речовиною. Так, поживна енергія, необхідна для функціонування клітини, надходить ззовні і витрачається здійснення нею різних функцій.
Структура найпростішої одиниці будови живого організму: мембрана органели, різноманітні включення. Вона оточена мембраною, всередині якої розташовується ядро та всі органели. Це мітохондрії, лізосоми, рибосоми, ендоплазматичний ретикулум. Кожен структурний елемент має власну мембрану.
Роль у життєдіяльності клітини
Біологічна мембрана відіграє кульмінаційну роль у будові та функціонуванні елементарної живої системи. Тільки клітина, оточена захисною оболонкою, може називатися організмом. Такий процес, як обмін речовин також здійснюється завдяки наявності мембрани. Якщо структурну цілісність її порушено, це призводить до зміни функціонального стану організму в цілому.
Клітинна мембрана та її функції
Вона відокремлює цитоплазму клітини від довкілля чи то з оболонки. Мембрана клітини забезпечує належне виконання специфічних функцій, специфіку міжклітинних контактів та імунних проявів, підтримує трансмембранну різницю електричного потенціалу. У ній є рецептори, здатні сприймати хімічні сигнали - гормони, медіатори та інші активні біологічні компоненти. Ці рецептори наділяють її ще однією здатністю – змінювати метаболічну активність клітини.
Функції мембрани:
1. Активне перенесення речовин.
2. Пасивне перенесення речовин:
2.1. Дифузія проста.
2.2. Перенесення через пори.
2.3. Транспорт, що здійснюється за рахунок дифузії переносника разом з мембранною речовиною або за допомогою передачі по естафеті речовини молекулярної ланцюга переносника.
3. Перенесення неелектролітів завдяки простій та полегшеній дифузії.
4. Активний транспорт іонів.
Будова мембрани клітини
Складові мембрани клітини - ліпіди та білки.
Ліпіди: фосфоліпіди, фосфатидилетаноламін, сфінгомієлін, фосфатидилінозит і фосфатиділсерін, гліколіпіди. Частка ліпідів становить 40-90%.
Білки: периферичні, інтегральні (глікопротеїни), спектрин, актин, цитоскелет.
Основний структурний елемент – подвійний шар фосфоліпідних молекул.
Покрівельна мембрана: визначення та типологія
Небагато статистики. На території Російської Федерації мембрана як покрівельний матеріал використовується не так вже й давно. Питома вага мембранних покрівель із загальної кількості м'яких перекриттів дахів становить лише 1,5 %. Більше поширення у Росії набули бітумні і мастичні покрівлі. А ось у Західній Європі частку мембранних покрівель припадає 87 %. Різниця відчутна.
Як правило, мембрана в ролі основного матеріалу при перекритті даху ідеально підходить для плоских покрівель. Для тих, хто має великий ухил, вона підходить меншою мірою.
Обсяги виробництва та реалізації мембранних покрівель на вітчизняному ринку мають позитивну тенденцію до зростання. Чому? Причини більш ніж зрозумілі:
- Термін експлуатації складає близько 60 років. Уявіть собі лише гарантійний термін використання, який встановлюється виробником, досягає 20 років.
- Легкість у монтажі. Для порівняння: монтаж бітумної покрівлі займає в 1,5 рази більше часу, ніж монтаж мембранного перекриття.
- Простота в обслуговуванні та проведенні ремонтних робіт.
Товщина покрівельних мембран може становити 0,8-2 мм, а середній показник ваги одного квадратного метра дорівнює 1,3 кг.
Властивості покрівельних мембран:
- еластичність;
- міцність;
- стійкість до впливу ультрафіолетових променів та інших середовищ-агресорів;
- морозостійкість;
- вогнетривкість.
Мембрана покрівельна буває трьох типів. Головний класифікаційний ознака - вид полімерного матеріалу, що становить основу полотна. Отже, покрівельні мембрани бувають:
- що належать групі ЕПДМ, виготовлені на основі полімеризованого етилен-пропілен-дієн-мономера, а простіше кажучи, Переваги: висока міцність, еластичність, водонепроникність, екологічність, низька вартість. Недоліки: клейова технологія з'єднання полотен за допомогою спеціальної стрічки, низькі показники міцності з'єднань. Сфера застосування: використовується як гідроізоляційний матеріал для тунельних перекриттів, водних джерел, сховищ відходів, штучних та природних водойм і т.д.
- ПВХ-мембрани. Це оболонки, при виробництві яких як основний матеріал використовується полівінілхлорид. Переваги: стійкість до ультрафіолету, вогнетривкість, велика кольорова гама мембранних полотен. Недоліки: низькі показники стійкості до бітумних матеріалів, олій, розчинників; виділяє в атмосферу шкідливі речовини; колір полотна з часом тьмяніє.
- ТПО. Виготовляються із термопластичних олефінів. Можуть бути армованими та неармованими. Перші оснащуються сіткою із поліестеру або скловолоконною тканиною. Переваги: екологічність, довговічність, висока еластичність, температуростійкість (як за високих, і за низьких температур), зварні з'єднання швів полотен. Недоліки: висока цінова категорія, відсутність виробників вітчизняному ринку.
Мембрана профільована: характеристика, функції та переваги
Профільовані мембрани – це інновація на будівельному ринку. Така мембрана експлуатується як гідроізоляційний матеріал.

Речовина, що використовується при виготовленні - поліетилен. Останній буває двох типів: поліетилен високого тиску (ПВД) та поліетилен низького тиску (ПНД).
Показник | ||
Міцність під час розриву (МРа) | ||
Подовження при розтягуванні (%) | ||
Щільність (кг/куб. м) | ||
Міцність при стисканні (МРа) | ||
Ударна в'язкість (з надрізом) (КДж/кв. м) | ||
Модуль пружності на вигин (МРа) | ||
Твердість (МРа) | ||
Робочий температурний режим (С) | від -60 до +80 | від -60 до +80 |
Добова норма водопоглинання (%) |
Профільована мембрана з поліетилену високого тиску має особливу поверхню - пустотілі пухирці. Висота цих утворень може коливатися від 7 до 20 мм. Внутрішня поверхня мембрани – рівна. Це дає можливість безпроблемного згинання будматеріалу.
Зміна форми окремих ділянок мембрани виключена, оскільки тиск по всій її площі розподіляється рівномірно завдяки наявності тих самих виступів. Геомембрана може використовуватися як вентиляційна ізоляція. У такому разі забезпечується вільний тепловий обмін усередині будівлі.
Переваги профільованих мембран:
- підвищена міцність;
- теплостійкість;
- стійкість хімічного та біологічного впливу;
- тривалий термін експлуатації (понад 50 років);
- простота в установці та обслуговуванні;
- доступна вартість.
Профільовані мембрани бувають трьох видів:
- з одношаровим полотном;
- із двошаровим полотном = геотекстиль + дренажна мембрана;
- із тришаровим полотном = слизька поверхня + геотекстиль + дренажна мембрана.
Одношарова профільована мембрана застосовується для захисту основної гідроізоляції, монтажу та демонтажу підготовки бетоном стін з підвищеною вологістю. Двошарову захисну використовують під час оснащення пристінного дренажу. Складається з трьох шарів застосовують на грунті, який піддається морозним пученням, і грунтовому грунті, що знаходиться глибоко.
Сфери використання дренажних мембран
Профільована мембрана знаходить своє застосування у таких областях:
- Основна гідроізоляція фундаменту. Забезпечує надійний захист від руйнівного впливу ґрунтових вод, кореневих систем рослин, осідання ґрунту, пошкоджень механічного типу.
- Стіновий дренаж фундаменту. Нейтралізує вплив грунтових вод, атмосферних опадів у вигляді переправлення в дренажні системи.
- Горизонтальний дренаж пластового типу – захист від деформації завдяки структурним особливостям.
- Аналог підготовки бетону. Експлуатується у разі проведення будівельних робіт зі зведення будівель у зоні низького залягання ґрунтових вод, у тих випадках, коли використовується горизонтальна гідроізоляція з метою захисту від капілярної вологи. Також функції мембрани профільованої входить непропускання цементного молока в грунт.
- Вентиляція стінових поверхонь підвищеного рівня вологості. Може встановлюватися як на внутрішній, так і зовнішній стороні приміщення. У першому випадку активізується повітряна циркуляція, а в другому забезпечується оптимальна вологість та температура.
- Інверсійна покрівля, що використовується.
Супердифузійна мембрана
Є матеріалом нового покоління, головним призначенням якого є захист елементів покрівельної конструкції від вітрових явищ, опадів, пари.

Виробництво захисного матеріалу ґрунтується на використанні нетканих речовин, щільних волокон високої якості. На вітчизняному ринку популярна тришарова та чотиришарова мембрана. Відгуки фахівців та споживачів підтверджують, що чим більше шарів лежить в основі конструкції, тим сильніші її захисні функції, а значить, і вища енергоефективність приміщення в цілому.
Залежно від типу даху, особливостей її конструкції, кліматичних умов, виробники рекомендують надавати перевагу тому чи іншому виду дифузійних мембран. Так, існують вони для скатних покрівель складних та простих конструкцій, для дахів скатного типу з мінімальним ухилом, для покрівель з фальцевим покриттям тощо.
Супердифузійна мембрана укладається безпосередньо на теплоізоляційний шар, настил із дощок. Необхідності у вентиляційному зазорі немає. Кріпиться матеріал спеціальними скобами або залізними цвяхами. Краї дифузійних листів з'єднуються роботи дозволяється проводити навіть за екстремальних умов: при сильних поривах вітру тощо.

Крім того, що розглядається покриття може використовуватися як тимчасове перекриття даху.
ПВХ-мембрани: сутність та призначення
ПФХ-мембрани - це матеріал для покрівлі, що виготовляється з полівінілхлориду і має еластичні властивості. Такий сучасний покрівельний матеріал витіснив бітумні рулонні аналоги, що мають істотний недолік - необхідність систематичного обслуговування і ремонту. Сьогодні характерні особливості ПВХ-мембран дозволяють використовувати їх при проведенні ремонтних робіт на старих покрівлях плоского типу. Застосовуються вони при монтажі нових дахів.

Покрівля з такого матеріалу зручна в експлуатації, а її встановлення можливе на будь-які типи поверхонь, у будь-яку пору року та за будь-яких погодних умов. ПВХ-мембрана має такі властивості:
- міцність;
- стійкість при впливі УФ-променів, різноманітних атмосферних опадів, точкових та поверхневих навантаженнях.
Саме завдяки своїм унікальним властивостям ПВХ-мембрани служитимуть вам вірою та правдою протягом багатьох років. Термін використання такої покрівлі прирівнюється до терміну експлуатації самої будівлі, тоді як рулонні покрівельні матеріали потребують регулярного ремонту, а в деяких випадках взагалі демонтажу та встановлення нового перекриття.
Між собою мембранні полотна з ПВХ з'єднуються методом зварювання гарячим зітханням, температура якого знаходиться в межах 400-600 градусів за Цельсієм. Така сполука є абсолютно герметичною.
Переваги ПВХ-мембран
Переваги їх очевидні:
- гнучкість покрівельної системи, що максимально відповідає будівельному проекту;
- міцний, з герметичними властивостями з'єднувальний шов між мембранними полотнами;
- ідеальна переносимість зміни клімату, погодних умов, температури, вологості;
- підвищена паропроникність, яка сприяє випару вологи, що накопичилася в підпокрівельному просторі;
- безліч варіантів колірних рішень;
- протипожежні властивості;
- здатність тривалий період зберігати первісні властивості та зовнішній вигляд;
- ПВХ-мембрана – абсолютно екологічний матеріал, що підтверджується відповідними сертифікатами;
- процес монтажу механізований, тому не триватиме багато часу;
- правила експлуатації допускають встановлення різних архітектурних доповнень безпосередньо зверху мембранної ПВХ-покрівлі;
- одношарове укладання заощадить ваші гроші;
- простота в обслуговуванні та ремонті.

Мембранна тканина
Текстильної промисловості мембранна тканина відома давно. З такого матеріалу виготовляється взуття та одяг: дорослий та дитячий. Мембрана - основа мембранної тканини, представлена у вигляді тонкої полімерної плівки і має такі характеристики, як водонепроникність і паропроникність. Для виробництва даного матеріалу цю плівку покривають зовнішнім та внутрішнім захисними шарами. Будова їх визначає сама мембрана. Це робиться з метою збереження всіх корисних властивостей навіть у разі пошкодження. Іншими словами, мембранний одяг не промокає при впливі опадів у вигляді снігу або дощу, але водночас добре пропускає пару від тіла у зовнішнє середовище. Така пропускна здатність дозволяє дихати шкірі.
Враховуючи все вищесказане, можна зробити висновок про те, що з подібної тканини виготовляється ідеальний зимовий одяг. Мембрана, що знаходиться в основі тканини, може бути:
- з порами;
- без пір;
- комбінована.

У складі мембран, що мають безліч мікропор, числиться тефлон. Розміри таких пір не досягають габаритів навіть краплі води, але більше водної молекули, що свідчить про водонепроникність та здатність виводити піт.
Мембрани, які не мають пір, як правило, виготовлені з поліуретану. Їхній внутрішній шар концентрує у собі всі потожирові виділення тіла людини і виштовхує їх назовні.
Будова комбінованої мембрани передбачає наявність двох шарів: пористого і гладкого. Така тканина має високі якісні характеристики і прослужить довгі роки.
Завдяки цим перевагам одяг та взуття, виготовлені з мембранних тканин і призначені для носіння в зимову пору року, міцні, але легкі, чудово захищають від морозу, вологи, пилу. Вони просто незамінні для багатьох активних видів зимового відпочинку, альпінізму.
Мембрани біологічні.
Термін "мембрана" (лат. membrana - шкірка, плівка) почали використовувати більше 100 років тому для позначення клітинного кордону, що служить, з одного боку, бар'єром між вмістом клітини та зовнішнім середовищем, а з іншого - напівпроникною перегородкою, через яку можуть проходити вода та деякі речовини. Однак цим функції мембрани не вичерпуються,оскільки біологічні мембрани становлять основу структурної організації клітини.
Будова мембрани. Відповідно до цієї моделі основної мембрани є ліпідний бислой, в якому гідрофобні хвости молекул звернені всередину, а гідрофільні головки назовні. Ліпіди представлені фосфоліпідпмі - похідними гліцерину або сфінгозину. З ліпідним шаром пов'язані білки. Інтегральні (транмембрані) білки пронизують мембрану наскрізь і міцно з нею пов'язані; переферичні не пронизують та пов'язані з мембраною менш міцно. Функції мембранних білків: підтримка структури мембран, отримання та перетворення сигналів з окр. середовища, транспорт деяких речовин, каталіз реакцій, що відбуваються на мембранах Товщина мембрани становить від 6 до 10 нм.
Властивості мембрани:
1. Плинність. Мембрана не є жорсткою структурою- більша частина білків і ліпідів, що входять до її складу, може переміщатися в площині мембран.
2. Асиметрія. Склад зовнішнього та внутрішнього шарів як білків, так і ліпідів різний. Крім того, плазматичні мембрани тварин клітин зовні мають шар глікопротеїнів (глікоколікс, що виконує сигнальну та рецепторні функції, а також має значення для об'єднання клітин у тканини)
3. Полярність. Зовнішня сторона мембрани несе позитивний заряд, а внутрішня негативний.
4. Виборча проникність. Мембрани живих клітин пропускають, крім води, лише певні молекули та іони розчинених речовин. іони розчинених речовин.)
Зовнішня клітинна мембрана (плазмалема) – ультрамікроскопічна плівка товщиною 7.5нм, що складається з білків, фосфоліпідів та води. Еластична плівка, що добре змочується водою і цілісність, що швидко відновлюється після пошкодження. Має універсальну будову, типову для всіх біологічних мембран. Прикордонне положення цієї мембрани, її участь у процесах виборчої проникності, піноцитоз, фагоцитоз, виведення продуктів виділення та синтез, у взаємозв'язку з сусідніми клітинами та захист клітин від пошкоджень робить її роль виключно важливою. Тварини зовні від мембрани іноді бувають покриті тонким шаром, що складається з полісахаридів і білків, - глікокаліксом. У рослинних клітин зовні від клітинної мембрани знаходиться міцна, що створює зовнішню опору та підтримує форму клітини клітинна стінка. Вона складається з клітковини (целюлози)-нерозчинного у воді полісахариду.





